The Use of GPS & GIS tools to help document the spread of desert mistletoe (Phoradendron californicum), a plant parasite in Arizona.
Juliann E. Aukema and Michael R. Kunzmann
Abstract:
Desert mistletoes are plant parasites dispersed by birds. The foraging patterns of seed-dispersing birds create seed shadows that with time, develop into mistletoe spatial distributions. The responses of seed-dispersers to resource abundance can create a positive feedback: high local abundances of mistletoes can lead to higher seed deposition. Using differential GPS, hosts GPS and GIS, Phoradendron californicum were mapped. Using a GIS database, layers of hosts, mistletoes dispersed seeds, and bird abundance were created. GIS can help us to understand and illustrate the spatial dynamics of pathogen transmission and spread.
Introduction:
The Santa Rita Experimental Range (SRER) (Figure 1), located at the foot of the Santa Rita Mountains about 35 miles south of Tucson in Pima County, Arizona, was established through a series of executive orders starting in 1900. By 1941, it had reached its present size of 53,159 acres. Today, SRER is under the jurisdiction of the USDA Forest Service and the University of Arizona and it is used to conduct research on a variety of aspects of semidesert rangeland ecology; and in addition to livestock grazing, the area is used for hunting and bird watching. Elevations range from 2900 feet in the northwest corner of the range to 5200 feet in the southeast. Simultaneously, average rainfall increases with elevation and ranges from 10 - 20 inches. SRER is home to 101 species of plants, 36 species of mammals, 46 species of birds, and 10 reptile species. Major changes in vegetation have been documented since the 1900's (Martin, 1986). For example, Prosopis velutina, velvet mesquite has become dominant on as much as 30,000 acres that were dominated by grassland 80 years ago (Medina, 1996). Information on the ecology of SRER is very comprehensive for a semidesert area of its size and diversity (Martin and Reynolds 1973 ).
Landscape and Ecological Interactions:
The desert mistletoe, Phoradendron californicum, (Figure 2), is a parasitic plant found in the deserts of northern México through Southern Utah. The desert mistletoe primarily parasitizes legume trees such as Mesquite (Prosopis spp.) (Figure 3), Acacia (Acacia spp.), Palo Verde (Cercidium spp.), and ironwood (Olneya spp.) (Figure 4). The desert mistletoe is dispersed by birds. Although quail, thrashers, mockingbirds, and woodpeckers consume mistletoe berries, the most effective disperser is Phainopepla nitens, (Larson, 1996) more commonly referred to by it's common name Phainopepla. The Phainopepla is the most northern of a group of tropical birds that feed on mistletoe. The Phainopepla is one member of the Silky Flycatcher family, Ptilogonatidae, that includes such tropical birds as the Gray Silky-flycatcher, Ptilogonys cinereus, found in Sonora, México, Phainoptila melanoxantha, the Black-and-yellow Silky-flycatcher, and Ptilogonys caudatus, the Long-tailed Silky-flycatcher, found in Panama, Costa Rica, and Guatemala.
Phainopeplas are easily recognized by their longish tail, crest, red eye, conspicuous upright posture, vocalizations, and treetop perching behavior. Adult birds are easily distinguished by differences in color (sexually dimorphic). Adult males are shinny black with contrasting white wing patches (Figure 5) while females and juveniles are gray with a pale-white wing patch (Figure 6). Typically they are known as desert scrub birds but may be found in a wide range of other habitats were mistletoe fruit is abundant throughout central California, Arizona (below the Colorado Plateau), southern Nevada, and west Texas.
Phainopeplas spend the winter in the desert, when flying insects are not abundant, and mistletoe berries become a primary food source, during the breeding season. Reportedly, phainopeplas consume at least 1100 mistletoe berries per day (Walsberg, 1975; Chu and Walsberg, 1999). During the summer, phainopeplas migrate from the deserts to higher elevations, where they breed again. In the desert these birds defend territories, whereas in the oak woodlands, they breed in loose colonies, feed together, and cooperate in mobbing potential predators such as scrub jays. The contrasting social system in a single species is unusual and is thought to be related to differences in the spatial distribution of food resources and nesting sites in the two habitats (Walsberg, 1977).
Phainopeplas have a specialized digestive system for consuming mistletoe berries. In the gizzard they remove the seed and pulp from the seed coat of the berries, they then digest the pulp and defecate the seeds and seed coat separately (Walsberg, 1975). When these seeds land on the branch of an appropriate host (Figure 7) they germinate and specialized penetration cells grow through the bark of the host to tap into the xylem, the tissue that transports water from the roots to the rest of the plant. The mistletoe absorbs water and minerals, for which it is entirely dependent on the host. But it is able to photosynthesize itself (hence it is a hemiparasite). In this system, phainopeplas are unusual because they have a dual role in the ecological community. They act as seed dispersers for the mistletoes and by dispersing these seeds, they also play the role of a disease vector, like a tick carrying Lyme disease or a mosquito transmitting malaria (Martínez del Rio et al., 1996).
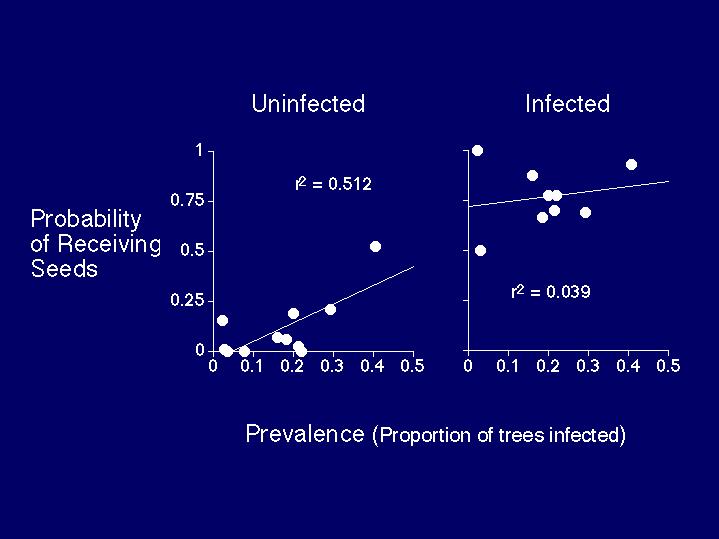
Because hosts, vectors, parasites, and infective propagules (seeds) are easily visible (Figure 8) mistletoes offer a unique opportunity to study pathogen transmission and seed dispersal in a spatial context. Seed-dispersing birds not only respond to their landscape, but can alter their landscape as well, creating a positive feedback. Preliminary field results (Aukema and Martínez del Rio, in prep.) have indicated that mistletoes are clumped within hosts - most hosts are not infected or have few parasites and a few hosts have most of the parasites (Figure 9) - and that the probability that a tree will have seeds deposited on it is positively correlated with the prevalence of mistletoes in its local neighborhood (Figure 10) (Aukema and Martínez del Rio, unpublished data). These field observations suggest that phainopeplas respond to mistletoes at the scale of individual trees and local neighborhoods and that mistletoes will be aggregated at larger spatial scales as well.
To examine the spatial distribution of mistletoes, hosts, phainopeplas, and concomitant seed rain, we mapped the locations of host trees and parasites on a four hectare plot in the northern part of the Santa Rita Experimental Range (Figure 11). A total of 729 individual trees were mapped representing 3 species: Prosopis velutina, Acacia greggii, and Cercidium microphyllum. In June , 1999, prior to GPS field activities to obtain coordinate sampling data, individual trees of the known host species were measured and tagged with a unique letter-number that would serve as a primary database key necessary for subsequent identification and to relate spatial coordinates to ecological attributes, and other landscape-level information. A survey was conducted in August and September 1999, to obtain the GPS location for each tree selected for this study.
To increase the spatial accuracy of all coordinate data and to reduce the errors associated with layer-to-layer registration and overlay analysis, all GPS data was "differentially" corrected to reduce typical GPS measurement errors, including but not limited to "selective availability" and atmospheric effects. A geodetic reference station (base-station) was established near the plot (Figure 12) to facilitate field work and reduce GPS errors associated with long-distances between the base station and the field GPS unit (rover). The rover was used to collect coordinate, ecological attributes, or landscape-level site information as deemed necessary in the field. Initial GPS-based database designs involved the collection of numerous field attributes such as tree-id, species, landscape feature, (x, y) offset, bearing, and other species characteristics to name a few. However, because of time and equipment power constraints, the number of mandatory attributes required at each location was reduced whenever feasible. The field objective was to move through the landscape quickly and efficiently while obtaining the coordinates on as many objects as possible.
To acquire spatial point data with 1-2 meter accuracy, a rover in kinematic mode, was carried to each tree to record attributes of interest such as the tree-id (Figure 13) For those trees whose canopy extended too far to get the GPS equipment quickly near or under the tree canopy (which may cause a loss of satellite signal lock), an offset distance was measured (Figure 14). Geolink (Baker-GeoResearch, Billings Montana) field mapping software was used as the computer interface to record attributes. By using Geolink software, offset accuracy could be visually checked by examining the relative position and bearing of points as they were captured and displayed on the screen.
In addition, as desired, computerized background maps (images) and GIS coverages could be examined in the context of a field data collection event. The integration of GPS data collection and attribution with GIS layer visualization in the field provides a greater opportunity review and comment on the combined spatial data. Typically GIS data are from numerous sources, often varying in spatial accuracy, and from differing time periods. This GPS-GIS in-field mapping technique creates an opportunity for new data discoveries and object-entity x, y relationships that may not be as obvious in the office or GIS laboratory.
Subsequent to field data collection intervals, the geodetic reference station was "post-processed" using two differential GPS software processing techniques - code acquisition (CA) and carrier phase - using two software packages Postpoint (Baker-GeoResearch Inc.) and Grafnav (Waypoint, Inc.) and all point locations were post processed using code acquisition.
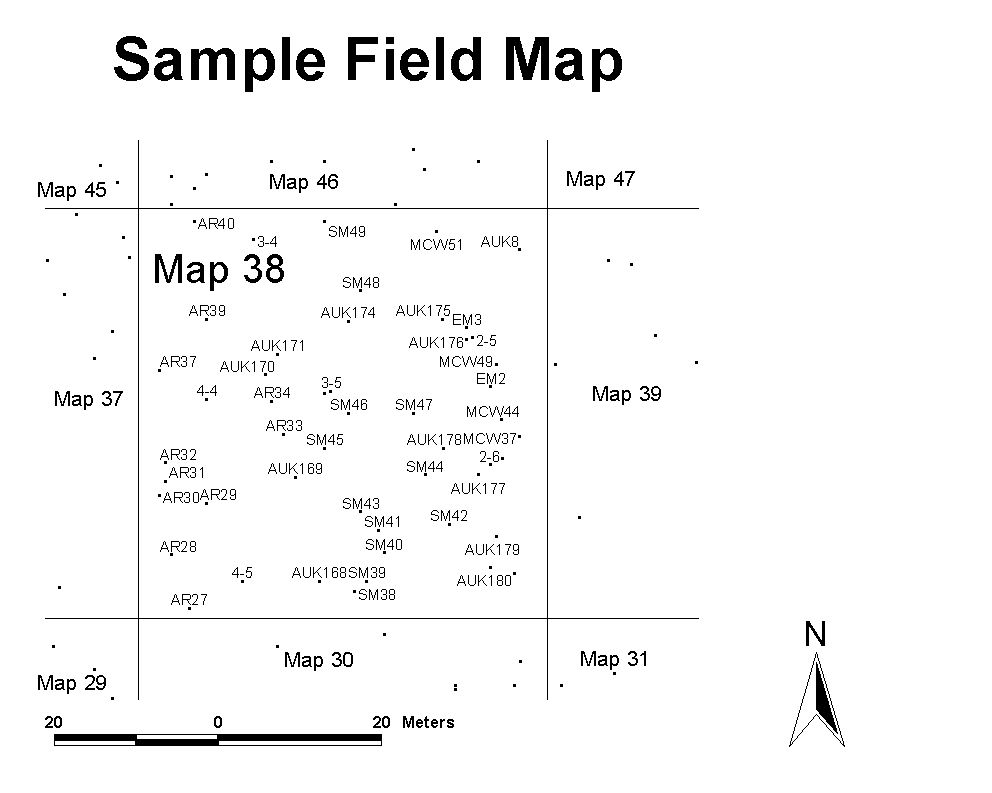
All the locations and attributes collected in the field (height, species, number of mistletoes etc.) were then joined using a combination of ArcInfo aml's and ArcView event tables. ArcView was used to generate views and map layouts and a map was generated. The points were ground truthed using a field map (Figure 15) when seeds were counted (Figure 16).
Discussion:
Using ArcInfo and ArcView we were able to produce a variety of maps that would give us some insight into the spatial distribution of the data. For example, species distribution maps (Figure 17) height class distributions (Figure 18), mistletoe distributions (Figure 19), and seed rain distributions (Figure 20) were easily generated from ArcView layouts. Maps are a useful way of visualizing the spatial distributions of host trees and mistletoes. However, only rigorous analysis can answer questions such as whether host of particular size classes are spatially autocorrelated, whether mistletoes are spatially autocorrelated, or whether the probability that a tree is parasitized or receives seeds is related to its neighborhood. Such analyses are difficult to do using "real world" data limited in duration to two field seasons.
After field data collection, integration with GIS software such as ArcInfo and a variety of descriptive and spatial statistical analysis packages frequently required reformatting of the data. It should be emphasized, that unless one is careful during data conversion process between as data is transported between software packages, errors may be generated that are not easily detected. Unfortunately, there is a lack of data quality assessment tools that actually check more than parity bits. Unanticipated problems we encountered as we moved data between vendors were numerous. For example, transferring data between an older version of PC ArcInfo (3.4.2) and more recent versions of ArcView (3.1) and workstation ArcInfo (7.0) resulted in a loss of decimal precision by data truncation.
Fortunately, this type of problem was easily recognized when the coverage was built and displayed (Figure 21). This type of error is also common with undefined spreadsheet templates and a wide range of other software that display and output different numbers of significant digits without notifying the operator.
There are also numerous, version (time) dependent, data conversion techniques between software suppliers that require time and effort to locate and learn. In addition, there are different database field structures, query processors, data dictionary and definition standards, cardinality mapping issues, and data aggregation differences that are not adequately defined. The primary point is that different data storage protocols should not produce different mathematical and analytical results without warning the operator that a spatial operation or calculation may result in the loss of data or precision. Furthermore it would be helpful if additional "standard" tools could be incorporated into software packages that would enhance spatial, analytical, and attribute manipulation capabilities.
One set of tools we would like to see integrated with ArcInfo would be the ability to statistically summarize tabular data diagonally, to produce omission/commission tables commonly used in remote sensing to determine classification accuracy. Spatial error tracking routines would also be helpful when layers or varying quality are aggregated to produce new results. Spatial errors often do not have normal distributions so non-parametric statistical routines should also be integrated with and or transparent to the GIS. Additional cooperation between software vendors, enhanced data translators, faster analysis tools, and better protocol and procedure documentation are required.
Conclusions:
GPS and GIS are powerful tools for examining the spatial distribution of plants and animals. This study, with the use of these tools, can give us valuable information about the distribution and spread of a plant parasite and about the effects of bird foraging behavior on the distribution of their resources and vice versa. One of the more important aspects of this study is finding the appropriate spatial scale at which to work. In addition, it is necessary to balance the need for resolution with the need to minimize sampling efforts. Future research needs to address patterns at scales larger than 4 hectares, patterns in communities with greater host species diversity, and spatial patterns of mistletoe demography.
Future research is required to make GIS and related spatial analysis tools more "user" friendly. Spatial sampling routines and protocols across multiple dimensions, datasets, and distance scales need to be developed. Universal query tools, data storage protocols, and data manipulation tools need to be standardized between major GIS and statistical software vendors. Spatial scale, resolution and sampling are everything! Fortunately, while not perfect, there has been significant technological progress toward seamless data integration and common standards.
Acknowledgements:
We would like to thank Carlos Martínez del Rio for help with methods, data collection, and analysis; Roy Hobbs for technical advice; John McCormack, Mike Saks, Rachel Boudin, Evan Canfield, the mapping crew, and Emma (Figure 22) for field help. We would like to thank the Advanced Resources and Technology lab at The University of Arizona for resources. This work was funded by a National Science Foundation graduate research fellowship, as well as by the Ecology and Evolutionary Biology department, RTG for the Analysis of Biological Diversification and Arid Lands department, all of the University of Arizona.
Chu, M. and G. Walsberg. 1999. Phainopepla (Phainopepla nitens). In: A. Poole and F. Gill, editors. The Birds of North America, No. 415. The Birds of North America, Inc., Philadelphia, PA.
Larson, D. 1996. Seed dispersal by specialist versus generalist foragers: the plant's perspective. Oikos 76:113-120.
Martin, S. C. 1986. Eighty years of vegetation change on a semidesert range in southern Arizona, U.S.A. and evaluation of causes. In: Ross, P. J.; Lynch, P. W.; Williams, O. B., eds. Rangelands: A resource under siege. Proceedings of Second International Rangeland Congress; 1986; Canberra, Australia. Sydney, Australia: Cambridge University Press: 53.
Martin, S. C. and H.G. Reynolds. 1973. The Santa Rita Experimental Range: Your facility for research on semidesert ecosystems. Journal of the Arizona Academy of Science. 8: 56-67.
Martínez del Rio, C., A. Silva, R. Medel, and M. Hourdequin. 1996. Seed dispersers as disease vectors: bird transmission of mistletoe seeds to host plants. Ecology 77: 912-921.
Medina, A. L. 1996. The Santa Rita Experimental Range: history and Annotated bibliography (1903-1988). General Technical Report RM-GTR-276. Rocky Mountain Forest and Range Experiment Station. Forest Service, U.S. Department of Agriculture.
Walsberg, G.E. 1975. Digestive adaptations of Phainopepla nitens associated with the eating of mistletoe berries. Condor 77:169-174.
Walsberg, G.E. 1977. Ecology and energetics of contrasting social systems in Phainopepla nitens (Aves: Ptilogonatidae). University of California Publications in Zoology 108: 1-63.
About the Authors:
Juliann Eve Aukema is a doctoral candidate in Ecology and Evolutionary Biology at the University of Arizona. Correspondence may be sent to Biological Sciences West 310, The University of Arizona, Tucson, Arizona, 85721. Juliann may also be reached by telephone at (520) 621-4022 or by email:jaukema@u.arizona.edu.
Michael R. Kunzmann is a USGS Ecologist with the USGS Sonoran Desert Field Station located at The University of Arizona, Tucson, Arizona. Correspondence may be sent to 125 Biological Sciences East, The University of Arizona, Tucson, Arizona, 85721. Mike may also be reached by telephone at (520) 621-7282 or by email: mrsk@sherpa.srnr.arizona.edu.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}