William W. Bushing
Geographic information systems have become established tools for analyzing species distributions in relation to environmental variables on land. This study utilizes a GIS to investigate the influence of terrestrial and submarine topography on the near-shore marine light regime, and the distribution of giant kelp (Macrocystis pyrifera), around an island off the southern California coast. Macrocystis is an important algal species along temperate coasts, serving both as structural habitat and food for many marine invertebrates and fishes. Santa Catalina Island's NW-to-SE orientation and highly dissected, 87-km leeward and windward coastline offers a wide range of microhabitats with respect to the nearshore light regime and factors such as exposure to winter storms and temperature.
GIS data layers included kelp distribution maps from several survey years, a digital terrain model, digital bathymetric model, submarine slope, submarine aspect and bottom relief. The topographic and bathymetric data layers were exported to Image Processing Workbench (IPW) to generate a series of integrated solar insolation models (monthly, seasonal, annual). An ocean surface insolation model incorporated the diurnal path of the sun, and the effect of hillshading on nearshore waters due to the adjacent island topography. Insolation on the sea floor added the elements of light extinction through the water column utilizing the bathymetric data and estimated regional turbidity. Kelp bed distribution was evaluated statistically relative to surface and bottom insolation levels.
Definite patterns in distribution were observed relative to surface and bottom insolation, especially pronounced during the peak growing season. As expected, submarine aspect, depth, slope, and bottom relief also played roles in influencing local light regimes. On the leeward side, shaded from mid-day and afternoon sun by the island's 450-640 meter main ridge, kelp favored the higher light environments around coastal features such as points and offshore reefs, or regions with greater bottom relief. Kelp on the more exposed, windward side of the island favored locations sheltered from the prevailing winter storms despite lower light levels in these sites.
As the energy source for several important ecological processes including photosynthesis, solar radiation is an important ecological parameter (Brock 1981, Nikolov and Zeller 1992). A knowledge of the spatial patterns of net surface radiation and its variability is important in understanding the growth and distribution of individual species, especially plants (Pearcy 1983, Duguay and Walker 1993); the distribution of plant and animal species within a community (Lakhani and Davis 1982, Weiss et al. 1988, Murphy and Weiss 1988); or ecosystem processes such as energy fixation and water balances (Lin et al. 1992. Hetrick et al. 1993a, 1993b).
Variability in light is influenced by a wide range of factors including Earth's rotation and orbit; atmospheric factors; surface albedo; and the surface topography. Significant variation in the light regime exists in regions with high relief due to variations in illumination angle and topographic shading (Dozier and Frew 1990). The importance of slope, aspect and elevation to the distribution of terrestrial plant species or communities has been noted by many authors (see Cantlon 1953, Yeaton and Cody 1979, Hicks and Franks 1984, Lieffers and Larkin-Lieffers 1987), and incorporated in models (see Geiger 1965 in Gates 1980, Holland and Steyn 1975, Temps and Coulson 1977, Holland et. al. 1977, Nunez 1980, Kirkpatrick and Hutchinson 1980). The effect of topographic shading is less well known (Hetrick et al. 1993b), but investigations of this factor are becoming more commonplace in the literature (Weiss et al. 1988, Murphy and Weiss 1988, Davis and Dozier 1990, Davis and Goetz 1990).
Light is also an important spatial variable in marine ecosystems, influencing the distribution of marine plants, invertebrates (Ebling et al. 1960 in Norton 1978; Hatcher 1989) and vertebrates (Frank and Leggett 1982). However, light has even greater variability in aquatic environments due to strong and selective absorption by water (Kirk 1983), differences in sediment loading and turbidity, bottom substrate type (Dean 1985), suspensoids (Kirk 1985), phytoplankton (Woods and Onken 1982, Pennock 1985, Attridge 1990), and even by-products released from marine plants like kelp (Lüning 1981a). Thom and Albright (1990) concluded that solar energy was the principal factor controlling seasonal physiological changes in benthic plants. Yet, this important variable has seldom been measured over extended periods and large spatial scales in the nearshore marine environment (see Kain 1982, Dean 1985). Undoubtedly this is due in part to the greater difficulty in modelling the light regimes of aquatic habitats.
Giant kelp (Macrocystis pyrifera) is a species of marine alga found along the Pacific coast of North America from central California to Baja California (Abbott and Hollenberg 1976). Although it begins life as a microscopic spore at the ocean floor, this species may grow to lengths of 60 m with its upper fronds forming a dense canopy at the surface. Giant kelp prefers depths less than 40 m, temperatures less than 20° C, hard substrate, and bottom light intensities above 1% that of the surface (North 1971, Foster and Schiel 1985). Macrocystis plays an important role in the marine environment by providing food and habitat for a wide range of marine invertebrates and fishes in southern California. Forests of giant kelp may support millions of individual organisms and more than 1,000 species of marine plants and animals (North 1971, Earle 1980, Foster and Schiel 1985).
Dean and Jacobsen (1984) felt that irradiance was the most important factor influencing growth in Macrocystis, especially during winter. Low light levels at specific locations or times may restrict giant kelp distribution by inhibiting sporophyte production as well as growth (Dean and Deysher 1983). Canopy-forming kelps like Macrocystis may also be light-limited during periods of fog or overcast (Jackson 1987, North et al. 1986). Several numerical models of kelp growth have also incorporated light as an important parameters (Jackson 1987, Nisbet and Bence 1989, Burgman and Gerard 1990).
North (1991) outlined a seasonal cycle in several physical factors influencing kelp distribution which included daylength. The ecological effects of varying daylength on marine plants due to season or latitude are well-known (see North 1971; Kirk 1983; Jackson 1987; Gonzalez-Fragosa et al. 1991; Lüning and Kadel 1993). What may not be as apparent is the influence of local topography in altering the effective photosynthetic daylength in the nearshore marine environment. Site-specific factors such as elevation, slope, aspect and the accompanying hillshading alter local light regimes creating marked spatial heterogeneity in incoming solar radiation. The seasonal (Kirkpatrick et al. 1988) or diurnal (Miklos et al. 1991) timing of peak insolation in regions of high relief may influence physiological or reproductive functions in a species.
Only a few studies, mostly terrestrial, have addressed the influence of topographic variability on regional diurnal and seasonal light regimes (see Dubayah et al. 1990). Although this has not been done for marine algae, Johansson (1982) found substantial differences in the species composition of freshwater algae between shaded and exposed locations. Seapy and Littler (1993) mention the role of afternoon shading by steep sea cliffs on neighboring Santa Barbara Island. Shading is becoming increasingly significant in modelling local light regimes as computational capacity increases (see Nunez 1980, Dozier and Frew 1990, Dubayah et al. 1990, Dubayah and Rich 1996).
Santa Catalina Island offers an excellent opportunity for investigation of such influences. Catalina is one of eight Channel Islands located off the southern California coast (figure 1). The island is in a tectonically active region with a dry climate, factors which contribute to the formation of high relief landscapes (Kirkpatrick et al. 1988). The island is oriented in a NW-to-SE direction (figure 2), has a high (450-600 m) central ridge and rugged topography. This orientation creates large-scale differences in insolation between the southwest-facing windward side and the northeast-facing leeward side. Afternoon sunlight intercepted by the island's main ridge reduces irradiance in the nearshore waters on the leeward side, effectively reducing daylength. In addition, the island's 54-mile coastline is highly dissected with small bays, points and other coastal features. This hillshading combined with the locally irregular coastline, variable submarine depths, a spatially heterogeneous atmosphere due to the maritime influence, and differential fog and cloudiness on the windward and leeward sides results in a wide range of microhabitats with respect to light and other environmental parameters. The island's steeper offshore slope and narrow shelf, compared to the adjacent mainland, force kelp closer to shore where it may be affected more by these variations in the light regime.
Previous island-wide studies using a geographic information system (GIS) have revealed significant differences in kelp distribution around the island due to differing disturbance regimes (temperature and storm), turbidity, submarine topography, and other factors (Bushing 1994, 1995). Field-measured data (Bushing 1996) indicates terrestrial hillshading reduces surface light levels to physiologically significant levels in the nearshore marine environment on the shaded leeward side. This study utilizes the same GIS coupled with a computer-generated surface insolation model to investigate the potential influence of topographic shading on the nearshore marine light regime, and the distribution of giant kelp around the island (figure 3)
In addition, light extinction in the water column as a function of depth and regional turbidity is incorporated to model spatial variability in the light regime on the ocean floor, and its potential influence on the younger life stages (gametophytes and young sporophytes) living there. Significant reductions in the nearshore light regime on the shaded leeward side should cause kelp to shift into unshaded, higher light environments around points, reefs and other coastal features, or further from shore where shading is less significant. Increased bottom relief creates a wider range of aspect and slope exposures, and thus potential light microclimates, along the ocean floor. This influence on kelp distribution will be tested using the insolation and bottom relief GIS layers. Finally, this information will be incorporated with the results from previous studies to generate guidelines for identifying kelp beds suitable for designation as future marine reserves.
GIS data layers required to address the hypotheses in this study include kelp bed distribution, terrestrial elevation, bathymetric depth, submarine aspect, submarine slope, bottom relief, distance-from-shoreline and solar insolation. The creation of the topographic (elevation, depth, aspect, slope and bottom relief) data layers has been described previously (Bushing 1995). The digital elevation model (DEM) and digital bathymetric model (DBM) were integrated to create a continuous topographic surface at 20-m resolution from a depth of 100 fathoms (183 m) to the highest peak, Mt. Orizaba (639 m) for use in modeling of the influence of local topography on incident light at the ocean surface and floor.
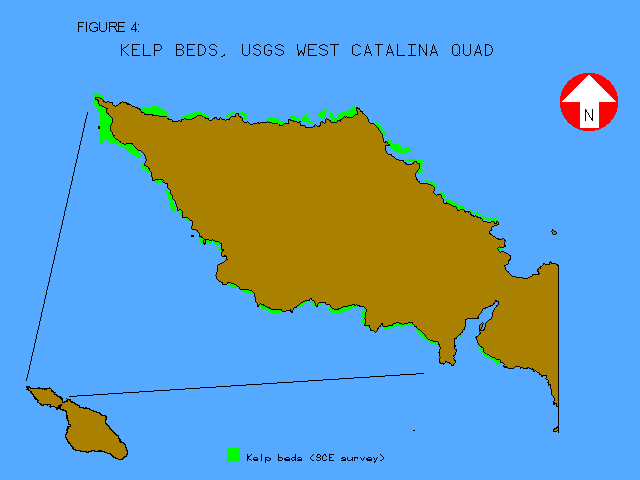
The distance-from-shoreline layer was created by generating the Euclidean distance of each ocean raster cell from the nearest island coastline. The initial kelp bed distribution map, dated about 1980 (figure 4), was provided by Southern California Edison in ArcInfo EXPORT format. Although several additional kelp distribution layers are available (Bushing 1995), only the 1980 data was analyzed relative to insolation in this study. All data layers were registered to base maps of the island's shoreline manually digitized from the four USGS 7.5' quad maps covering Santa Catalina Island at a scale of 1:24,000 using UTM-11 coordinates.
To investigate the influence of island topography on the light regime required GIS layers to model the effects of terrestrial hillshading on the nearshore marine environment, and light extinction in the water column. Previously, Bushing (1994, 1995) modeled instantaneous light levels at the ocean surface and floor within a GIS. Those results indicated differences between the windward and leeward coasts in kelp distribution relative to insolation. However, this simple model represented only a single time slice in the diurnal and seasonal variability of the sun's path across the sky and its interplay with the island's topography. To better understand the influence of light, and topographic effects, a model integrating insolation over time periods of days, months, seasons or annually is required.
A large body of literature, and many different light models, exist for habitats in the terrestrial environment. Several authors offer general reviews of the factors involved in applying insolation models (see Okanoue 1957; Okanoue and Makijama 1958; Robinson 1966; Revfiem 1978, 1982; Duffie and Beckman 1980; Badescu 1981; Ideriah 1981). Others address the modelling of light specifically in regions of high relief (Nunez 1980; Dozier 1980, 1989; Dubayah 1992; Hetrick et al. 1993a, 1993b). Several solar insolation models are available within commercial GIS software including the HILLSHADE (see Dubayah and Rich 1996) and SOLARFLUX (see Rich et al. 1993; Hetrick et al. 1993a, 1993b; Saving et al. 1993; Weiss et al. 1993; all in Dubayah and Rich 1996) models in Esri's ArcInfo and GRID software.
For this study the TOPQUAD module, developed by Dozier (1980, 1989) for integration into the IPW (Image Processing Workbench) software toolkit (Frew 1990), was chosen. The TOPQUAD algorithm, its precursors and derivatives have been used extensively for modeling light environments in regions of high relief (see Dozier 1980, Dozier et al. 1981, Frew and Dozier 1986, Davis and Dozier 1990, Davis and Goetz 1990, Dozier and Frew 1990, Dubayah et al. 1990, Dubayah and Rich 1996). The increased ability to parameterize the TOPQUAD algorithm, allowing it to more closely model local light regimes and overall heterogeneity, coupled with the ease of transporting images from IPW software to various GIS packages, made TOPQUAD an appropriate choice for this study.
For physiological or ecological studies only certain regions of the electromagnetic spectrum may be important (McCree 1972, Brock 1981, Dubayah and Rich 1996). For this study, only the wavelengths (400-700 nm) utilized in photosynthesis, known as PAR (photosynthetically active radiation), were used. Insolation was modeled for the 21st day of each month to create monthly intervals between solstices and equinoxes. Similar methods were used by Weiss et al. (1988), Murphy and Weiss (1988) and Hetrick et al. (1993a), although other methods have also been suggested (Klein 1976, Duffie and Beckman 1980, Dubayah et al. 1990, Davis and Goetz 1990, Lieffers and Larkin-Lieffers 1987). Because insolation in successive months tends to be highly correlated, values were composited in three month intervals by averaging image pixels in IPW to create "seasonal" images. October, November and December were averaged to create a "fall" insolation composite (see figure 5, figure 6a and figure 6-b). This allowed correlation with the seasonal aspects of the Macrocystis growth and life cycles. All twelve monthly models were also averaged to create an "annual" insolation model.
FIGURE 5: TOPQUAD solar insolation model for the fall months, West quad, Santa Catalina Island
To incorporate the effect of regional turbidity and light extinction with depth on young kelp at the ocean floor where the species must first establish, a GIS layer was generated from the TOPQUAD surface insolation layer to model bottom insolation. The model assumes the deepest depth for Macrocystis occurs at a value of 1% of surface irradiance (PAR), the generally accepted limit of the photic zone (Lüning and Dring 1976; Lüning 1981a; Kirk 1983, 1985; Ramus 1983, Walsh, et al. 1987, Smith et al. 1987). Relative turbidity was estimated by determining the maximum depth (dmax) at which kelp was found on the windward and leeward coasts.
The extinction of light through any medium is defined by Beer's Law (Gates
1980, Kirk 1983, Jones et al. 1983, Jackson 1987). Based on this
relationship, the index representing insolation at the ocean floor was generated
using image algebra to multiply the surface insolation value (Is) by a light
extinction equation for each image location as follows:
where bottom insolation (Id) is the percent of surface insolation (Is) from the first model at that cell's depth (d), and dmax is the maximum depth kelp was found in the geographic region. This light extinction equation was applied cell-by-cell, using the depth in each cell, to create seasonal and annual bottom insolation values incorporating the effects of the diurnal and seasonal solar arc, terrestrial hillshading, light extinction through the water column, and regional turbidity. Both the Is and Id indices assume clear skies and do not account for local variations in cloud cover or fog.
Insolation is strongly dependent on aspect (Boyko 1947, Holland and Steyn 1975). Kelp frequency distributions generated for 10° intervals (eg, 30-40°) of aspect, and polar graphs of mean surface and bottom insolation relative to aspect were prepared to analyze this relationship. Based on the frequency distributions, "typical" and "atypical" aspects were defined for each coast based on an arbitrary cut off of 5% frequency. Those aspects with frequency values greater than 5% were designated "typical" (0-60° and 330-360° on the leeward coast; 180-270° on the windward side). Aspects with frequency less than 5% were designated "atypical" (61-329° on the leeward coast; 0-179° and 271-360° on the windward side). Previous studies (Bushing 1994, 1995) indicate the "atypical" aspects generally have higher percent cover of kelp than "typical" aspects. The frequency distribution, percent cover of the photic shelf, and mean values with standard errors for both kelp and the photic shelf ("ocean") were also plotted as a function of 30° aspect intervals (eg, 30-60°) for each of the four seasonal, and the annual, surface and bottom insolation models.
Increased bottom relief creates a wider range of aspect and slope exposures, and thus potential light microclimates, along the ocean floor. Because bottom relief exhibited marked differences relative to aspect on both coasts (Bushing 1994, 1995), surface and bottom insolation was investigated relative to this variable. Mean surface and bottom insolation for kelp were calculated and graphed as a function of bottom relief.
To test the hypothesis that greater observed distances from shore along the leeward coast were a response to terrestrial hillshading, graphs were prepared to show the relationship between winter insolation as a function of distance from shore. Mean winter surface and bottom insolation were calculated and graphed as a function of distance from shore in 25 m intervals. Then distance was treated as the dependent variable, and mean distance from shoreline was calculated for 10 unit intervals of annual surface insolation.
Many of the data layers were stored both as polygon and raster data to facilitate the analyses in this study at differing spatial scales. Although insolation models for the entire island were generated, only the West windward and leeward coasts were analyzed in this study due to the data processing needs associated with multiple models. Although analysis was conducted using the vector polygon layers, much of the information presented here looked at the data on a cell-by-cell basis due to the influence of smaller-scale (~20 m) heterogeneity on kelp distribution. Binary raster masks representing kelp distribution from the 1980 SCE survey were created at 20 m resolution from the original polygon data for both leeward and windward coasts. The 20 fathom (~40 m) bathymetric contour was chosen to define the maximum expected depth limit for kelp. The region between this contour and the shoreline represents the potential area that might be occupied by kelp. For this study it is referred to as the "photic shelf" since its lower limit is generally considered to be light-limited. A second set of binary masks corresponding to the "photic shelf" on each coast was created using the shoreline and 20 fathom contours.
Coincident data was extracted from the raster GIS layers for submarine aspect, depth, slope, bottom relief, distance-from-shoreline, surface and bottom insolation using the kelp and photic shelf binary masks and overlay techniques. This data was analyzed within the GIS and the resulting grids and binary masks were converted to ASCII format for import into Foxpro databases representing the windward and leeward sides. Each data layer (eg, binary kelp distribution mask, extracted aspect) was imported as a separate field in the database so each record represented a single geographic location (raster cell) incorporating the associated data in multiple fields. Several programs were written in the xBASE computer language to further analyze the data on a cell-by-cell basis.
Two important statistics were evaluated for each data layer: kelp frequency and kelp percent cover. In this paper the phrase "kelp frequency" refers to the percentage of total kelp along each coastline with a given data value or range (eg, 5% of all kelp on the West leeward coast was found at surface insolation values of 150 to 160). The phrase "percent cover" refers to the percent of the corresponding "photic shelf" covered by kelp for a given value of an environmental variable (eg, 20% of all 8 m depths on the West windward coast were covered by kelp).
All hypotheses were tested in the following manner. The null hypothesis is that there are no significant topographic effects on the distribution of kelp relative to insolation on either coast. It is tested by comparing the observed kelp distributions against the values expected using the corresponding photic shelf distribution as a probability model (Johnson et al. 1988). The expected probabilities were calculated by multiplying the total number of kelp cells times the observed frequency (%) for the corresponding shelf region for each 10 unit range of the corresponding insolation model index (eg, 210-220 summer insolation). Residual values were obtained by subtracting the observed values for kelp from the expected value. Divergences from that distribution, as measured by the residuals, indicate parameter values which are preferred (+) or avoided (-). Graphs of kelp frequency distributions and calculated residual values with respect to each of the five solar insolation models were generated. If the two distributions do not differ in a statistically significant way, insolation and topographic effects are not factors in determining the distribution of kelp. If significant differences exist, topographic influences on light may play a role in effecting Macrocystis distribution. Data was also imported from the relational databases into statistics packages (SYSTAT version 5.02 and S-PLUS version 3.2) for further analysis and graphical representation (Wilkinson et al. 1992, Statistical Sciences Inc. 1993). Additional statistical methods were based on those of Bishop (1983).
In prior work (Bushing 1994), GIS-based modelling revealed that instantaneous late afternoon surface light levels for the island's leeward side were significantly lower (p < 0.001) than those for the windward side. Later field measurements (Bushing 1996) indicated that at leeward sites affected by terrestrial hillshading, incident sunlight was reduced to levels 1-6% that of adjacent exposed locations. In addition to reductions in instantaneous light levels, effective daylength was also reduced by 30% at the hillshaded sites compared to adjacent sites directly exposed. These results strongly suggest that the spatial patterns in the nearshore light regime introduced by hillshading may have physiologically significant effects on a short-term basis.
Much of the statistical analysis for the TOPQUAD model discussed here is presented elsewhere (Bushing 1996). In that report, sites where kelp was present had consistently, and significantly (p < 0.050), higher mean insolation values for the annual and 4 seasonal models than regions where kelp was not found on the leeward coast. However, light levels on the windward side were significantly lower (p < 0.001) in all but the fall and winter models for regions with kelp.
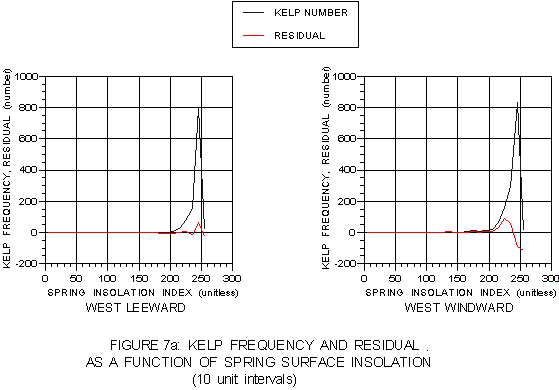
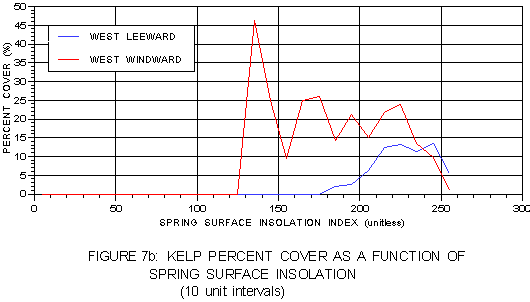
These relationships are easier to visualize when kelp distributions relative to surface and bottom insolation are presented graphically. Figure 7a presents kelp frequency and calculated residual values as a function of the spring surface insolation index. Kelp frequency tends to peak towards the upper end of the insolation range on both coasts. However, the calculated residual values show kelp frequency tends to exceed expected values in different portions of the two respective ranges. Residual values peak at the lower end of the windward side range, suggesting more kelp is present in regions of lower light than the probability model predicts, most likely representing nearshore sites. The leeward side exhibits a contrasting trend with kelp exceeding expectations toward the mid- or high-end of the range, most likely representing less-shaded offshore sites. Kelp percent cover as a function of spring surface insolation is presented in figure 7b. As expected based on the residual analysis, kelp on the leeward side shows peak percent cover at the upper end of its range, but windward kelp peaks at the lower end.
Due to the island's orientation, kelp frequency relative to aspect is dominated by the prevailing coastal aspects on the windward and leeward coasts (Bushing 1996). For the West leeward coast, "typical" aspects generally face NNW-to-NE (330-360°, 0-60°) versus "atypical" aspects (61-329°), while on the West windward side "typical" aspects face S-to-W (180- 269°) versus "atypical" (0-179°, 270-360°) aspects. Only 17.8% of total kelp on the leeward side was found at "atypical" aspects while the value was substantially higher (27.3%) for the windward coast.
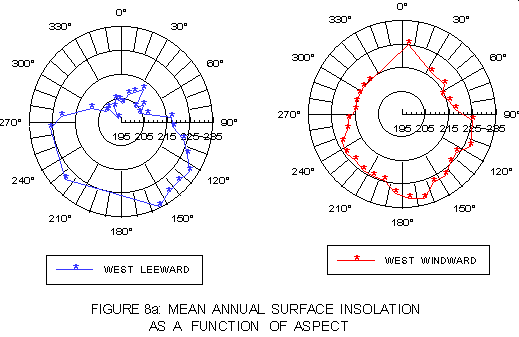
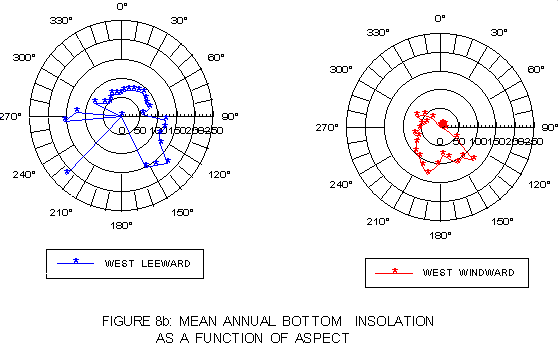
The strong influence of aspect on the light regime is illustrated in graphs of mean annual surface (figure 8a) and bottom (figure 8b) insolation as a function of 10° intervals of aspect. Mean leeward coast surface insolation is greater at "atypical" than "typical" aspects, but generally lower on the windward side. With respect to bottom insolation, kelp on the leeward side shows substantially higher light levels at "atypical" than at "typical" aspect values, while windward side values are lower at "atypical" aspects.
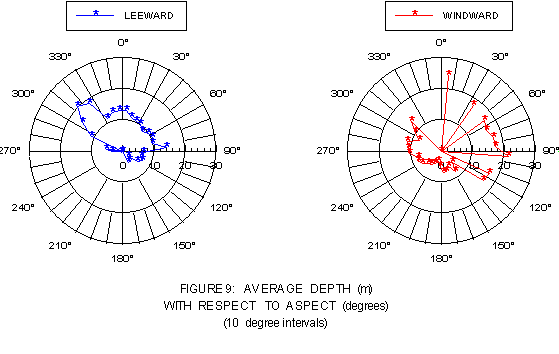
The difference in bottom light levels between "typical" and "atypical" aspects is partially explained by plotting mean depth as a function of aspect for the leeward and windward sides (figure 9). On both coasts, kelp is found at deeper depths at "atypical" aspects, but the difference is not statistically significant (p > 0.050) on the leeward side, but is (p < 0.001) on the windward side. Average depths on the windward side were generally less than 10-15 m for aspects from about 150° to 315°, but noticeably deeper (12-25 m) for aspects of 30° to 100°. Leeward side values averaged less than 12-15 m at aspects of 330° to 150°, but 15-35 m at aspects of 300° to 330°.
FIGURE 9: Mean depth as a function of aspect (10° intervals) for the windward and leeward coasts
"Atypical" aspects often represent locations around coastal points, offshore reefs or other features where the irregularity of the coastline provides a wider range of exposures. On the more protected leeward side, in the absence of severe storm disturbance, kelp may favor sunnier sites at "atypical" aspects around such features. The lower light levels at "atypical" aspects on the windward side suggest protection from storm exposure may override light as a controlling factor (Bushing 1995) on a coastline where light levels are generally high, but winter storm disturbance more significant.
Mean seasonal and annual surface and bottom insolation levels, and the statistical significance of any differences between "typical" and "atypical" aspects were previously reported (Bushing 1996). For the leeward coast, surface and bottom insolation was significantly higher (p < 0.050 or greater) at atypical aspects for the year, and most seasons except spring. On the windward side, surface insolation values tended to be lower at atypical aspects for all seasons, while bottom insolation values were significantly lower (p << 0.001) at atypical aspects. Again, this is consistent with light being a controlling factor on the leeward, and storm exposure on the windward sides.
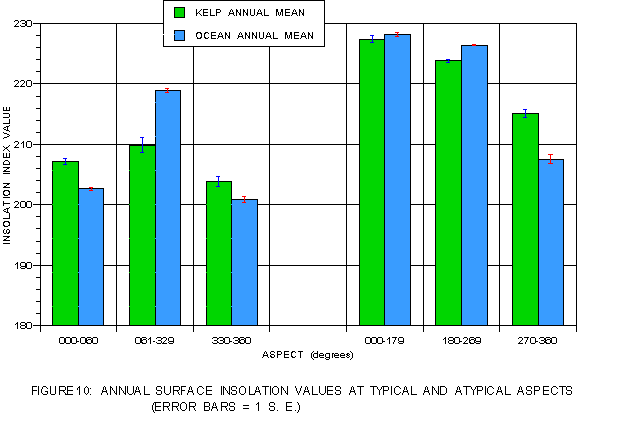
Figure 10 graphically illustrates the relationship between surface insolation for regions occupied by kelp and the entire photic shelf ("ocean") at the typical and atypical aspects previously defined for both coasts. Kelp at the typical leeward aspects (0-60° and 330-360°) exhibits higher mean values than the corresponding photic shelf, indicating a preference for sites with higher insolation. Values for kelp at atypical aspects (61-329°) are lower than for the photic shelf, but are still higher than the values at typical aspects. Since these aspects are interpreted to represent sunnier sites near coastal features, it is probable that the higher photic shelf insolation values represent regions away from the coast and therefore unshaded, but in deeper water where kelp may be unable to establish.
On the windward side, surface insolation values tend to follow the expected pattern based on exposure to the sun. The photic shelf at east-facing (0-179°) atypical aspects, which receive more sun during the fall and winter months, tends to have the highest surface insolation followed by the southwest-facing (180-269°) typical aspects and the northwest-facing (270-360°) atypical aspects, which receive less sunlight during those periods. However, surface insolation values for kelp at typical aspects (180-269°) are lower than for the corresponding photic shelf. Kelp at the northwest-facing (270-360°) aspects uniformly have higher values than the corresponding photic shelf, indicating they are favoring sunnier locations here where surface insolation values are lower than elsewhere on this coast.
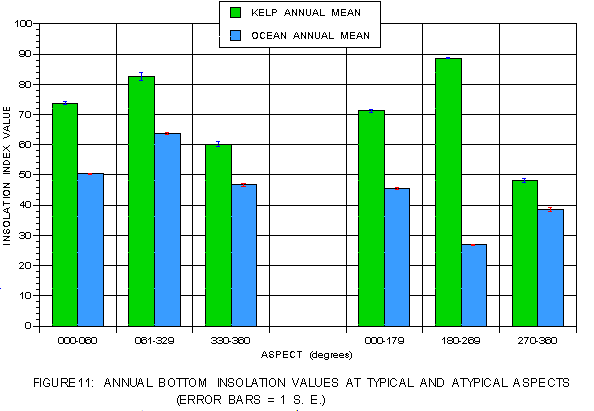
Bottom insolation (figure 11) indicates that on both coasts kelp values always substantially exceed those for the corresponding photic shelves at all aspect ranges. On the leeward side, the bottom light values for kelp at atypical aspects always exceed those for the typical aspect ranges regardless of sun exposure. On the windward side, the reverse is true with light values at the atypical aspects lower than those for the typical aspects. Distribution is optimized for bottom insolation on the leeward, but not the windward side.
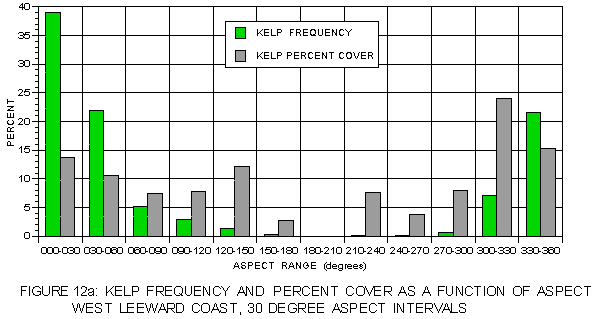
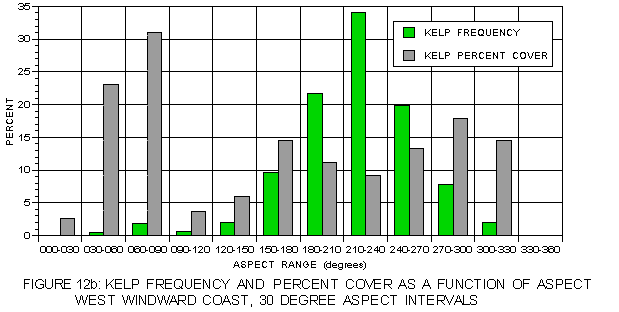
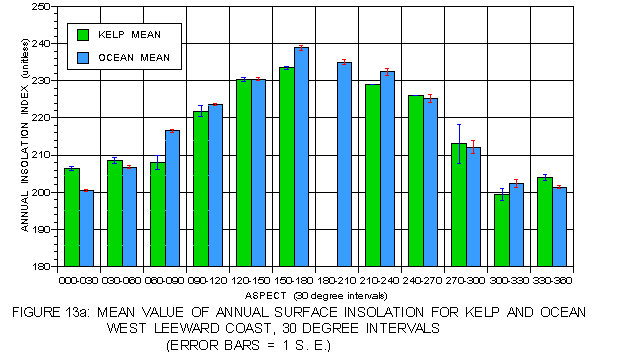
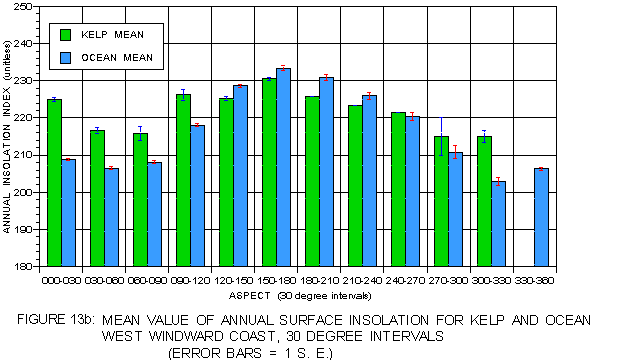
Even higher resolution in the pattern of light as a function of aspect was obtained by breaking aspect into 30° ranges. Kelp frequency and percent cover for each of these aspect ranges is illustrated in figure 12a and figure 12b. The nearly Gaussian frequency distributions are apparent in these figures, as is the generally higher percent cover at aspects with lower frequency. Annual surface insolation (figure 13a) on the leeward side exhibits a pattern in which values are strongly dependent on solar exposure. The highest values are at southeast-facing aspects (150-180°) and the lowest at northwest aspects (300-360°). Mean insolation values for kelp at the typical (330-60°) aspects exceed those for the corresponding photic shelf suggesting a preference for sites with higher surface light values. At atypical aspects, mean values for kelp are generally less than for the photic shelf, but these values are substantially higher than those at the typical aspects. The pattern on the windward coast (figure 13b) is different. For aspects of 240 to 120° kelp values exceed those of the corresponding photic shelf, but are lower at aspects from 120-240° where sunlight is strongest. This suggests some preference for sites with higher insolation where hillshading is a factor, but no preference in well-lit locations.
FIGURE 12a: Kelp frequency at aspect intervals of 30° on both coasts
FIGURE 12b: Kelp percent cover at aspect intervals of 30° on both coasts
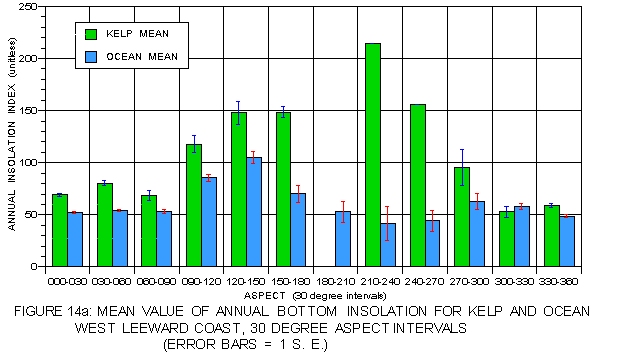
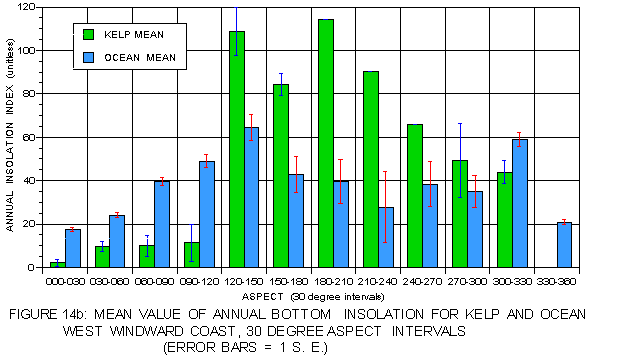
The patterns for annual bottom insolation on the leeward (figure 14a) and windward (figure 14b) coasts are quite different from that of surface insolation. Kelp values are substantially higher than those for the corresponding photic shelves on both coasts, reflecting an avoidance of the deepest depths with high light extinction. On the leeward coast, mean values for kelp are much higher than for the photic shelf at atypical than typical aspects while on the windward side the reverse is true. This pattern is consistent with that observed in all other analyses. Kelp on both sides favors atypical aspects but for different reasons. On the protected, but shaded leeward side these locations offer higher insolation values. Atypical aspects on the exposed, but well-lit windward side have lower light levels but provide protection from storm exposure, a more important factor there.
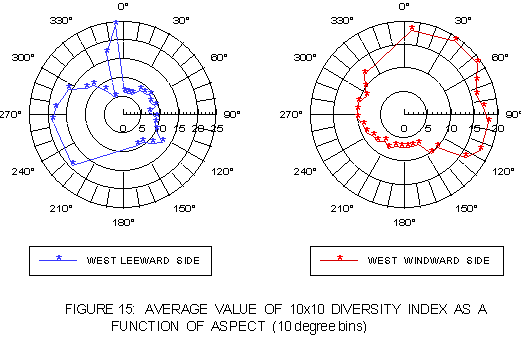
Previously kelp was shown to favor regions of greater bottom relief (Bushing 1995, 1996). Additional information may be gained by looking at the distribution of insolation relative to bottom relief. Figure 15 depicts the mean value for bottom relief in the kelp distributions as a function of 10° aspect intervals (eg, 20-30°). On both windward and leeward coasts, bottom relief tends to be higher at the atypical aspects than the typical aspects for kelp. This suggests atypical aspects include a greater proportion of points or offshore reefs.
Figure 16 illustrates the mean winter surface insolation values as a function of bottom relief. On the leeward side there is a trend towards increasing insolation with increasing bottom relief, consistent with the interpretation that kelp favors high relief sites where light levels may be greater due to more favorable sun exposure. On the windward side, winter surface insolation reveals a slight decline with increasing bottom relief. This suggests light levels may be lower in high relief regions, consistent with the hypothesis that atypical aspects on this coast provide protection from storm disturbance rather than advantageous light environments.
FIGURE 16: Mean winter surface insolation as a function of bottom relief on both coasts
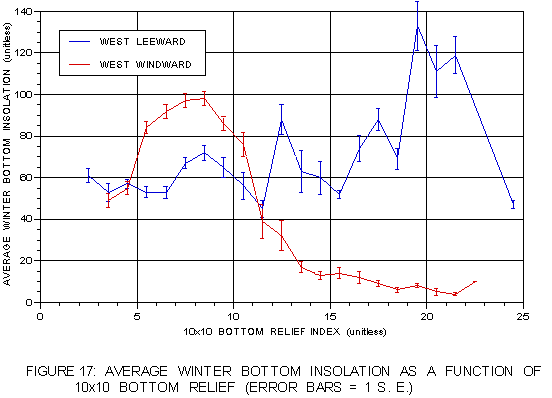
The winter bottom insolation pattern (figure 17) is more revealing. The same pattern of increasing insolation with increasing bottom relief is evident on the leeward side. However, on the windward side insolation shows a more complex trend. Insolation values peak at levels of moderate bottom relief (5-10), and steadily decline as bottom relief increases after that. One interpretation is locations of moderate bottom relief, such as offshore reefs, offer favorable environments with higher bottom insolation. In areas of high relief, near coastal points, the biological trade-off may be increased protection from storms despite lower insolation levels.
FIGURE 17: Mean winter bottom insolation as a function of bottom relief on both coasts
As previously reported (Bushing 1994) the mean distance from shoreline was significantly greater on the leeward (91.0 m) than the windward side (66.7 m), but the maximum distances were comparable at 331 m and 340 m respectively. Horizontal displacement of kelp away from shore on the leeward side, due to hillshading by the island's main ridge, may be responsible.
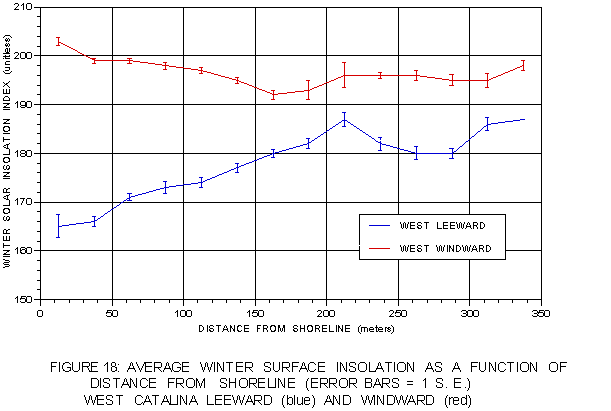
Figure 18 graphically depicts the relationship between mean winter surface insolation and distance from shoreline. On the leeward side, there is a pronounced increase in average surface insolation away from the coast to a distance of about 220 m. Within this distance, the horizontal displacement of kelp away from the coast may be due to hillshading. Beyond that distance, the pattern is irregular, suggesting offshore reefs play a role at these distances. On the windward side there is a slight decrease in average surface insolation with increasing distance from shore.
FIGURE 18: Mean winter surface insolation as a function of distance from shoreline on both coasts
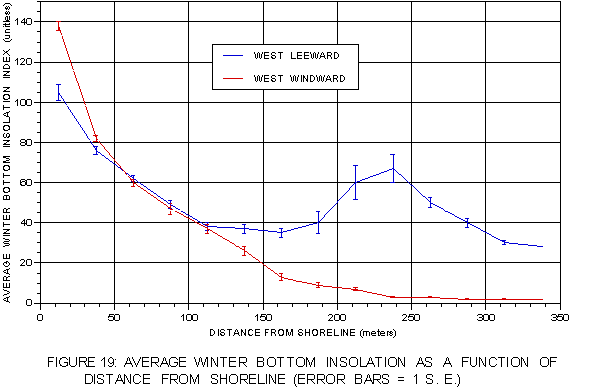
Average bottom insolation values as a function of distance from shore (figure 19) support this interpretation. On the windward side, average insolation drops exponentially as distance increases, reflecting increasing depth with distance. On the leeward side the drop is less precipitous, especially at distances from about 100 to 200 m from shore. The unexpected peak in bottom insolation values at distances of 200-300 m appears to confirm the influence of offshore reefs on the light environment at greater distances.
FIGURE 19: Mean winter bottom insolation as a function of distance from shoreline on both coasts
Based on field measurements and the GIS insolation model, the instantaneous light levels and effective daylength in the nearshore marine environment surrounding Catalina Island exhibit an extremely high degree of spatial variability due to seasonal changes, topography (slope and aspect), terrestrial hillshading, and regional differences in turbidity and overcast. These differences appear to influence the distribution of giant kelp around the island. The relationships observed between insolation and kelp distribution at "typical" and "atypical" aspects mirrors findings in the terrestrial environment (eg, Boyko 1947, Holland and Steyn 1975, Holbo and Childs 1987, Youtie et al. 1988, Allen et al. 1991). While the GIS values exhibit statistical significance, due in part to the large sample sizes, it must also be determined if they are physiologically or ecologically significant. This requires the incorporation of measured regional surface insolation from other studies, and the evaluation of light requirements for compensation, saturation, gametogenesis and other processes in giant kelp.
Mature kelp canopies absorb sunlight at its highest intensity near the surface, providing the adult sporophytes with relative immunity from changes in light due to absorption in the water column. Dense Macrocystis canopies may reduce surface light up to 99% (Neushul 1971, Pearse and Hines 1979, Kimura 1980, Reed and Foster 1984, North et al. 1986). The younger stages at the ocean floor experience diminished, and potentially wide fluctuations in, bottom insolation (Heine 1983, North et al. 1986).
Macrocystis must recruit and grow as a gametophyte or young sporophyte on the ocean floor, a relatively high nutrient environment where irradiance may be limiting, whereas the adult plants have most of their tissue in the canopy near the water surface where irradiance is high and nutrients may be low (Wheeler 1978; Dean and Jacobsen 1984, 1990). Individual plants must establish in the first regime before they can grow large enough to benefit from near-surface light conditions. The light requirements of gametophytes, as the stage responsible for successful recruitment, may be far more critical for the ecological success and persistence of kelp due to the large range of light regimes experienced at the bottom (Rosenthal et al. 1974, Harger 1979, Reed and Foster 1984, Deysher and Dean 1984).
The literature reports a range of field- and lab-measured light requirements for compensation, saturation and gametogenesis in different Macrocystis life stages. Critical light levels for giant kelp's growth and survival have been variously reported in the 40-220 µE m-2 sec-1 range (Wheeler 1978, Lüning 1981b, Smith et al. 1983, Gerard 1984, Zimmerman and Robertson 1985, North et al. 1986). The minimum level for gametophyte photosynthetic compensation or vegetative growth has been reported in the 2.8-20 µE m-2 s-1 (Lüning and Neushul 1978, Fain and Murray 1982, Deysher and Dean 1984) based on typical surface irradiance of 2,500 µE m-2 s-1, with higher levels required for the production of gametes. Some of this variability may stem from interactions between other variables such as temperature or nutrients (Dean and Deysher 1983, Deysher and Dean 1984). It is quite possible that the light requirements for kelp on Catalina's windward and leeward sides may differ due to differences in their temperature and nutrient regimes.
Regional surface insolation values have been reported by several authors (see Bennett 1965 in Gates 1980, Wheeler 1978, Harger 1979, Matta and Chapman 1991) and bottom light levels by others (Deysher and Dean 1984, 1986; Dean and Jacobsen 1984, Dean 1985). Wheeler (1978) reported winter and summer noon surface light levels of 1,926 and 3,853 µE m-2 s-1 at latitudes of about 30° N, comparable to Catalina. Fain and Murray (1982) reported typical surface irradiance levels of 2,500 µE m-2 s-1. Values from the clear sky TOPQUAD model indicate surface insolation over the photic shelf in winter, when variance is highest, vary by nearly 20:1. Field data indicate terrestrial hillshading may cause real world irradiance values to vary by a factor of 30:1 between adjacent sites, and reduce effective daylength by 40% or more. When light extinction and turbidity are incorporated, variation in bottom insolation exceeds 1,000:1, resulting in real world light levels below compensation range for kelp in this model under clear skies.
Harger (1979) found off Santa Barbara that noon bottom light levels were sufficient for gametogenesis in all months at least part of the time, although he suggested that light levels for the triggering of gametogenesis probably occurs rarely in winter and most often in spring or summer. Based on the model in this study, kelp on both windward and leeward coasts appear to optimize distribution relative to spring insolation. The overall high incident light values for southern California due to latitude and weather patterns may compensate somewhat for the effects of turbidity on the exposed windward coast and hillshading on the leeward side.
The ability of a plant to adapt to a given light regime depends on its genetic flexibility (Wheeler 1978). Some plants found in lower light environments, such as shaded habitats or at deeper depths, have a higher photosynthetic efficiency than members of the same species from sunny or shallow water habitats (Bjorkman 1973, Boardman 1977, Wheeler 1978, Lüning and Neushul 1978, Heine 1983). The observed presence of Macrocystis in deeper water on the shaded leeward side may be explained by such enhanced efficiency. Such light ecotypes are known from other marine algae (Gerard 1988, 1990) where higher photosynthetic efficiencies in plants from turbid waters were attributed to their greater nitrogen uptake capacity. It is interesting to note Kopczak et al. (1991) found Macrocystis off Catalina's shaded leeward side also was more efficient in its nutrient uptake and utilization.
If Macrocystis ecotypes adapted to different light regimes exist, this could have consequences for the selection of a marine reserve network around Catalina Island. Gerard (1988) suggested the limited gene flow between kelp populations due to short spore dispersal distances might maintain the geographic isolation of such ecotypes. Individuals adapted to the higher turbidity environments of the windward side would most likely incur a higher metabolic cost involved in maintaining higher pigment concentrations. This could be difficult under the more oligotrophic conditions that prevail on the leeward side of the island.
The extinction of light in the water column significantly influences the potential habitat for marine vegetation. Light often determines the maximum survival depth for marine algae at a given site (Lüning 1971, Lüning and Dring 1979, Gerard 1988), and is believed to limit the seaward extent of Macrocystis pyrifera, especially in turbid coastal waters (North 1971, Mel 1977, Neushul Mariculture Inc. 1981).
The physical aspects of submarine illumination and extinction through the water column have been discussed by Jerlov (1951, 1968, 1976), Tyler and Smith (1970), Gates (1980), Kirk (1983), and others. Its relevance to marine algae is reviewed in Lüning (1981a), Jones et al. (1983), Dean (1985) and Jackson (1987). Lüning (1981a) shows the percentage of downward irradiance (350-700 nm) in relation to depth in the different Jerlov water types. Based on his graph, and, the apparent depth limit (dmax) of the nearshore photic zone from the GIS, the waters off the West windward coast would be Jerlov type 5 and those off the leeward side Jerlov type 3. These values are comparable to the range observed by Dean (1985) off the mainland coast near San Onofre.
Light extinction and submarine illumination may be modified by a range of different factors: the water molecules themselves, dissolved substances, and particulate matter including suspended sediment, detritus and phytoplankton. These factors are often seasonal, such as the resuspension of sediments by winter storms (Neushul et al. 1976, Wheeler 1978) or phytoplankton blooms in late spring.
Seasonal and diurnal fluctuations in surface irradiance are generally not a major factor in the variability of bottom irradiance levels compared to changes in water clarity (Harger 1979, Heine 1983, Dean and Jacobsen 1984, Dean 1985, Dunton 1990). Dean (1985) found that variations in sediment levels related to storm activity, rather than seasonal surface irradiance, was responsible for observed fluctuations in bottom insolation in mainland southern California kelp beds. Bottom irradiance ranged from 0.1-11% of surface levels, a factor of 100X indicating a highly variable light regime at the ocean bottom.
Seasonal fluctuations in turbidity due to increased winter storm activity may be especially important if surface insolation is simultaneously reduced by low sun angles and increased hillshading, higher rainfall and accompanying sediment loading from terrestrial runoff, or higher levels of overcast. Regional differences in seasonal bottom insolation between the windward and leeward coasts off Catalina may be greater due to differing seasonal storm regime and wave energy. The windward side experiences sediment loading not only from runoff, but also from resuspension of sediments in the high energy environment. Reduced winter surface light levels on the leeward side due to topographic hillshading may somewhat balance this.
Although light intensity has been the primary focus of this model, spectral quality is also altered through selective extinction in the water column and may influence photosynthesis or other light-dependent processes (Kirk 1983). In coastal waters, where phytoplankton and organic substances are present, blue light is attenuated more strongly (Craigie and McLachlan 1964, Sieburth and Jensen 1968, Lüning 1981a, Kirk 1983). Higher coastal turbidity on the windward side or phytoplankton blooms may affect Macrocystis gametogenesis, which is dependent on exposure to blue light (Lüning and Neushul 1978).
The results in this study must be viewed in terms of the limitations of, and any associated error imposed by, the GIS model. Dubayah and Rich (1996) review such sources of error, identifying the poor quality of digital elevation models as most important. Others (Dozier 1990, Davis and Dozier 1990, Dubayah et al. 1990, Dubayah and van Katwijk 1992) emphasize the influence of resolution in the terrain model (DEM). Errors in this important dataset are propagated through each step in the modelling process since many of the elements (aspect, slope, view factors) used in the calculations are derived from the original topographic data layer. The optimum resolution, or grid spacing, in DEM's used for ecological modeling of light fields depends on the geomorphology of the region, and the ecologically relevant spatial scales of interest. Regions of high relief like Catalina require DEM's with higher spatial resolution to more accurately model fine-scale topographic variation.
Dozier (1980), Choudhury (1982) and Dubayah (1992) emphasized the importance of surface albedo in modeling the spatial distribution of radiation. Although this study assumed a uniform value, the TOPQUAD model permits cell-by-cell parameterization of surface albedo in estimating the surface-reflected component. Dubayah and Rich (1996) state that the average albedo is usually sufficient, yet terrestrial vegetation and soil types in high relief regions may differ substantially. The vegetation on the east-facing leeward slopes is often dense scrub or chaparral, which is less reflective than the barren, sparsely-vegetated windward slopes, further exaggerating the differences due to hillshading. A good vegetation map, recoded with albedo values appropriate to each vegetation type, or satellite imagery might improve the accuracy of this model (Sjoberg and Horn 1983, Dubayah 1992).
The TOPQUAD model may fail in nearshore marine environments like that off Catalina due to the significant difference in the nature of terrestrial (Lambertian) and water surfaces (specular) as reflectors. Differences in wave activity on the windward and leeward sides may also affect modeled bottom light levels since the albedo of the sea surface is also a function of solar angle (Holmes 1957, Attridge 1990, Wing et al. 1993, Taylor and Stephens 1983).
The results of this study indicate that the light regimes, as well as effective daylengths, at the ocean surface and floor in the nearshore marine environment surrounding Santa Catalina Island exhibit an extremely high degree of spatial variability due to seasonal changes, the influence of terrestrial hillshading, and regional differences in turbidity. This spatial heterogeneity appears to be physiologically significant, and to have influenced the distribution of Macrocystis pyrifera around the island.
This study offers the first evidence of the potential influence of terrestrial hillshading on the distribution of Macrocystis pyrifera. Hillshading reduces light values and effective daylength in the nearshore waters of some regions on the leeward side. Kelp distribution there favors sites with higher-than-average surface and bottom insolation, generally at atypical coastal aspects, while on the windward side distribution is more uniform with respect to light. The consistently larger kelp beds on the windward side (Bushing 1994) may also be due in part to longer daylength, higher light and photosynthetic levels.
Kelp's greater average distance from the coast on the leeward side reinforces the interpretation that hillshading plays a factor in influencing distribution. Such horizontal displacement may allow the interception of more light for photosynthesis by the adult sporophyte canopy at the surface. On the leeward side Macrocystis is also associated with regions of moderate to high relief, representing coastal points and offshore reefs where light levels tend to be higher. On the windward side light levels tend to be higher in regions of moderate relief (reefs) rather than high relief (coastal features) which represent refugia from storms there. Direct exposure to prevailing wind, swell and storm patterns on this coast have generated higher levels of turbidity and increased light extinction in the nearshore waters (Bushing 1994, 1995), influencing the depth distribution of kelp around the island.
Reduced growth levels due to lower light levels on the shaded leeward side, or at protected aspects on the windward side, may result in higher mortality and diminished ability to resist the effects of the prevailing disturbance regime on each coast (Steinman et al. 1991). If so, regional persistence in kelp may be affected by the observed spatial variability in the regional light regime (Bushing 1995). Those individuals, ecotypes and populations which are able to survive may respond once the light regime increases due to removal of adult canopy by storm, reduced turbidity or other seasonal change. Resilience and persistence should be higher in such populations. Such spatial differentiation in local population characteristics should be incorporated as a factor in the identification of potential new marine reserves around the island.
The island's unique geography and topography offer an interesting test case. However, the model should also be extended to new geographic locations including other Channel Islands with differing geographic orientations, as well as mainland locations where hillshading may also be a factor around coastal points and promontories.

The author would like to acknowledge the following: the Catalina Island Conservancy and the Offield Family Foundation for their support in the development of several elements in this GIS; UCSB's Map & Imagery Lab, especially Larry Carver, Greg Hajic and Mark Probert; Esri and Statistical Sciences, Inc. for their kind donations of software to the Catalina Island Conservancy which helped make the project possible; Southern California Edison for the contribution of a kelp data layer in ArcInfo format; SPOT Image Corporation and Dr. Thomas Belsher of IFREMER (France) for the SPOT satellite imagery; Dr. M. Neushul, Dr. A. Ebeling, Dr. L. Mertes, Dr. J. Markham, Dr. D. Reed and Dr. K. Lafferty for their encouragement and advice along the way; Dr. J. Dozier for his suggestion of the TOPQUAD algorithm; Janet Takara for her monumental patience; and many other "anonymous" participants in various INTERNET mail lists and news forums, including Esri-L, GIS-L and comp.infosystems.gis who, unwittingly and unacknowledged, helped through their advice.
Abbott, I. and G. Hollenberg 1976. Marine Algae of California. Stanford University Press. Stanford, California. 827 pp.
Allen, R. B., R. K. Peet and W. L. Baker, 1991. Gradient analysis of latitudinal variation in southern Rocky Mountain forests. Jour. of Biogeography 18(2):123-139.
Attridge, T. H., 1990. Light and Plant Responses. Edward Arnold (New York). 148 pp. Badescu, V., 1981. A verification of the atmospheric model proposed by Barbaro et al. for computing direct and diffuse solar radiation. Solar Energy 26(5):459-460.
Badescu, V., 1981. A verification of the atmospheric model proposed by Barbaro et al. for computing direct and diffuse solar radiation. Solar Energy 26(5):459-460.
Bishop, O. N., 1983. Statistics for Biology. Longman House (Essex, England). 232 pp.
Bjorkman, O., 1973. Comparative studies on photosynthesis in higher plants. pp. 1-63. in A. C. Geise (ed). Photophysiology, vol. 5.
Boardman, N. K., 1977. Comparative photosynthesis of sun and shade plants. Ann. Rev. Plan Physiol. 28:355-377.
Boyko, H. 1947. On the role of plants as quantitative climate indicators and the geo-ecological law of distribution. Jour. Ecology 35:138-157.
Brock, T. D., 1981. Calculating solar radiation for ecological studies. Ecological Modelling 14(1-2):1-19.
Burgman, M. A. and V. A. Gerard, 1990. A stage-structured, stochastic population model for the giant kelp Macrocystis pyrifera. Marine Biology 105:15-23.
Bushing, W. W., 1994. The influence of topography on the distribution of giant kelp (Macrocystis pyrifera) beds around Santa Catalina Island using a geographic information system. In W. L. Halvorson and G. J. Maender (eds.), The Fourth California Islands Symposium: Update on the Status of Resources. Santa Barbara Museum of Natural History Santa Barbara, CA). pp. 70-82.
Bushing, W. W., 1995. Identifying regions of persistent giant kelp (Macrocystis pyrifera) around Santa Catalina Island for designation as marine reserves. Proc. 15th Esri User Conference, Palm Springs, CA, May 21-26, 1995. Proceedings published on CD-ROM and via World Wide Web (URL: http://www.Esri.com).
Bushing, W. W., 1996. The use of satellite remote sensing and a geographic information system (GIS) to characterize regions of persistent giant kelp (Macrocystis pyrifera) around Santa Catalina Island for designation as marine reserves. Unpublished PhD dissertation. University of California, Santa Barbara.
Cantlon, J. E., 1953. Vegetation and microclimate on north and south slopes of Cashetunk Mountain, New Jersey. Ecol. Monogr. 23:241-270.
Choudhury, B., 1982. A parameterized model for global insolation under partially cloudy skies. Solar Energy 29(6):479- 486.
Craigie, J. S. and J. McLachlan 1964. Excretion of coloured ultraviolet-absorbing substances by marine algae. Can. J. Bot. 42:23- 33.
Davis. F. W. and J. Dozier, 1990. Information analysis of a spatial database for ecological land classification. Photogr. Engr. and Remote Sensing 56(5):605-613.
Davis, F. W. and S. Goetz, 1990. Modeling vegetation pattern using digital terrain data. Landscape ecology 4(1):69-80.
Dean, T. A., 1985. The temporal and spatial distribution of underwater quantum irradiation in a southern California kelp forest. Estuarine, Coastal and Shelf Science 21(6):835-844.
Dean, T. A. and L. E. Deysher 1983. The effects of suspended solids and thermal discharges on kelp Macrocystis. In Bascom, W. (ed.), The Effects of Waste Water Discharges on Kelp in Southern California. Southern California Coastal Water Research Project, Long Beach, California. pp. 114-135.
Dean, T. A. and F. R. Jacobsen, 1984. Growth of juvenile Macrocystis pyrifera (Laminariales) in relation to environmental factors. Marine Biology 83(3):301-311.
Dean, T. A. and F. R. Jacobsen, 1990. Nutrient-limited growth of juvenile kelp, Macrocystis pyrifera, during the 1982- 1984 "El Niño" in southern California. Marine Biology 90:597-601.
Deysher, L. E. and T. A. Dean, 1984. Critical irradiance levels and the interactive effects of quantum irradiance and dose on gametogenesis in the giant kelp, Macrocystis pyrifera. Jour. Phycology 20(4):520-524.
Deysher, L. E. and T. A. Dean, 1986. In situ recruitment of sporophytes of the giant kelp, Macrocystis pyrifera (L.) C. A. Agardh: effects of physical factors. J. Exp. Mar. Biol. Ecol. 103:41-63.
Dozier, J. 1980. A clear-sky spectral solar radiation model for snow-covered mountainous terrain. Water Resources Research 16(4):709-718.
Dozier, J., 1989. On-line computer documentation for the contributed TOPQUAD module in the IPW (Image Processing Workbench) dated July 6, 1989.
Dozier, J., J. Bruno and P. Downey, 1981. A faster solution to the horizon problem. Computers & Geosciences 7:145- 151.
Dozier, J. and J. Frew, 1990. Rapid calculation of terrain parameters for radiation modeling from digital elevation data. IEEE Trans. on Geoscience and Remote Sensing 28(5):963-969.
Dubayah, R., 1992. Estimating net solar radiation using Landsat Thematic Mapper and digital elevation data. Water Resources Res. 28(9):2469-2484.
Dubayah, R. and P. M. Rich, 1996. GIS-based solar radiation modeling. In Goodchild, M. F., L. T. Steyaert, B. O. Parks, C. Johnston, D. Maidment, M. Crane and S. Glendinning (eds.). GIS and Environmental Modeling: Progress and Research Issues. Proc. 2nd Intl. Conference/Workshop on Integrating Geographic Information Systems and Environmental Modeling. Sept. 26-30, 1993. Breckenridge CO. Natl. Center for Geographic Information and Analysis. GIS World Books (Fort Collins, CO), pp. 129-134.
Dubayah, R. and V. van Katwijk, 1992. The topographic distribution of annual incoming solar radiation in the Rio Grande River Basin. Geophysical Research Letters 19:2231-2234.
Dubayah, R., J. Dozier and F. W. Davis, 1990. Topographic distribution of clear-sky radiation over the Konza Prairie, Kansas. Water Resources Research 26(4):679-690.
Duffie, J. A. and W. A. Beckman, 1980. Solar Engineering of Thermal Processes. Wiley-Interscience (New York, NY). 762 pp.
Duguay, C. R. and D. A. Walker, 1993. Environmental modeling and monitoring at the Niwot LTER site: an overview of recent research activities using GIS. In Proc. Second Intl. Conference/Workshop on Integrating Geographic Information Systems and Environmental Modeling, Sept. 26-30, 1993. Breckenridge, CO. Natl. Center for Geographic Information and Analysis. no page numbers.
Dunton, K. H., 1990. Growth and production in Laminaria solidungula: relation to continuous underwater light levels in the Alaskan High Arctic. Marine Biology 106(2):297-304.
Earle, S. 1980. Undersea World of a Kelp Forest. National Geographic, September, 1980. pp 411-426.
Ebling, F. J., Sleigh, M. A., Sloane, J. F. and J. A. Kitching 1960. The ecology of Lough Ine. VII. Distribution of some common plants and animals of the littoral and shallow sublittoral regions. Jour. Ecology 48:29-53.
Fain, S. and S. Murray, 1982. Effects of light and temperature on net photosynthesis and dark respiration of gametophytes and embryonic sporophytes of Macrocystis pyrifera. Jour. Phycol. 18:92-98.
Foster, M. S. and D. Schiel, 1985. The ecology of giant kelp forests in California: a community profile. U. S. Fish and Wildlife Service Biological Report 85(7.2). National Coastal Ecosystems Team. Slidell, LA. 152 pp.
Frank, K. T. and W. C. Leggett, 1982. Environmental regulation of growth rate, efficiency, and swimming performance in larval capelin (Mallotus villosus), and its application to the match/mismatch hypothesis. Can. Jour. Fish. Aquat. Sci. 39(5):691-699.
Frew, J. and J. Dozier 1986. The Image Processing Workbench- portable software for remote sensing instruction and research. Proceedings of the 1986 International Geoscience and Remote Sensing Symposium, European Space Agency, Paris, pp. 271-276.
Frew, J. E., 1990. The Image Processing Workbench. Ph.D. Dissertation. Dept. of Geography, University of California at Santa Barbara. 305 pp.
Gates, D. M., 1980. Biophysical Ecology. Springer-Verlag (New York) Geiger, R. 1965. The Climate Near the Ground. Harvard Univ. Press (Cambridge, MA).
Gerard, V. A., 1984. Physiological effects of El Ni¤o on giant kelp in southern California. Mar. Biol. Lett. 5:317-322.
Gerard, V. A., 1988. Ecotypic differentiation in light-related traits of the kelp Laminaria saccharina. Marine Biology 97(11):25-36.
Gerard, V. A., 1990. Ecotypic differentiation in the kelp Laminaria saccharina: phase-specific adaptation in a complex life cycle. Marine Biology 107(3):519-528.
Gonzalez-Fragosa, J., S. E. Ibarra-Obando and W. J. North, 1991. Frond elongation rates of shallow water Macrocystis pyrifera (L.) Ag. in northern Baja California, Mexico. Jour. of Applied Phycology 3(4):311-318.
Harger, B. W. W., 1979. Coastal Oceanography and Hard Substrate Ecology in a Californian Kelp Forest. PhD Dissertation. Department of Biology, University of California at Santa Barbara. 427 pp.
Hatcher, A., 1989. Variation in the components of benthic community structure in a coastal lagoon as a function of spatial scale. Austr. Jour. Mar. Freshwater Res. 40:79-96.
Heine, J. N., 1983. Seasonal productivity of two red algae in a central California kelp forest. Jour. Phycology 19:146-152.
Hetrick, W. A., P. M. Rich, F. J. Barnes and S. B. Weiss, 1993a. GIS-based solar radiation flux models. Proc. ASPRS 1993 Annual Conference, New Orleans, 3:132-143.
Hetrick, W. A., P. M. Rich and S. B. Weiss, 1993b. Modeling insolation on complex surfaces. In Proc. 13th Annual Esri User Conference. Environmental Systems Research Institute (Redlands, CA). pp 447-458.
Hicks, R. R. and P. S. Franks, 1984. Relationship of aspect to soil nutrients, species importance and biomass in a forested watershed in West Virginia. For. Ecol. Mgmt. 8:281-291.
Holbo, H. R. and S. W. Childs, 1987. Summertime radiation balances of clearcut and shelterwood slopes in southwest Oregon. Forest Science 33(2):504-516.
Holland, P. G. and D. G. Steyn, 1975. Vegetational responses to latitudinal variations in slope angle and aspect. Journal of Biogeography 2:179-183.
Holland, P. G., D. G. Steyn and R. F. Fuggle 1977. Habitat occupation by Aloe ferox Mill. (Liliaceae) in relation to topographic variations in direct beam solar radiation income. Journal of Biogeography 7:61-72.
Holmes, R. W. 1957. Solar radiation, submarine daylight and photosynthesis. In Hedgpeth, J. W. (ed.) Treatise on Marine Ecology and Paleoecology, Volume 1, Ecology. Geological Society of America, Memoir 67. pp. 109-28.
Ideriah, F. J. K., 1981. A model for calculating direct and diffuse solar radiation. Solar Energy 26(5):447-452.
Jackson, G. A., 1987. Modelling the growth and harvest yield of the giant kelp Macrocystis pyrifera. Marine Biology 95(4):611-624.
Jerlov, N. G. 1951. Optical studies of ocean water. Rep. Swedish Deep-Sea Exped. 3:1-59.
Jerlov, N. G. 1968. Optical Oceanography. Elsevier Publ. (New York). 194 pp.
Jerlov, N. G. 1976. Marine Optics. Elsevier Publ. (Amsterdam). 231 pp.
Johansson, C., 1982. Attached algal vegetation in running waters of Jämtland, Sweden. Acta Phytogeographica, suecica 71:12-73.
Johnson, L. B., C. A. Johnston and J. Pastor, 1988. Raster and vector data in ecological research applications. In Proceedings Third Annual International GIS/LIS '88 Conference, November 30 - December 2, 1988. pp. 386-94.
Jones, R. C., K. Walti and M. S. Adams, 1983. Phytoplankton as a factor in the decline of the submersed macrophyte Myriophyllum spicatum L. in Lake Winagra, Wisconsin, U.S.A. Hydrobiologia 107(3):213-219.
Kain, J. M., 1982. Morphology and growth of the giant kelp Macrocystis pyrifera in New Zealand and California. Marine Biology 67:143-157.
Kimura, S. 1980. The effects of harvesting Macrocystis pyrifera on the understory algae in Carmel Bay, California. M. S. Thesis, California State University, Fresno, CA. 108 pp.
Kirk, J. T. O., 1983. Light and Photosynthesis in Aquatic Ecosystems. Cambridge University Press (New York). 401 pp.
Kirk, J. T. O., 1985. Effects of suspensoids (turbidity) on penetration of solar radiation in aquatic ecosystems. Hydrobiologia 125:195-208.
Kirkpatrick, J. B. and C. F. Hutchinson, 1980. The environmental relationships of California coastal sage scrub and some of its component communities and species. Journal of Biogeography 7:23-38.
Kirkpatrick, J. B., R. J. Fensham, M. Nunez and D. M. J. S. Bowman, 1988. Vegetation-radiation relationships in the wet-dry tropics: granite hills in northern Australia. Vegetatio 76(3):103-112.
Klein, S. A., 1976. Calculation of monthly average insolation on tilted surfaces. Solar Energy 19:325.
Kopczak, C. D., R. C. Zimmerman and J. N. Kremer, 1991. Variation in nitrogen physiology and growth among geographically isolated populations of the giant kelp, Macrocystis pyrifera (Phaeophyta). Jour. Phycology 27(2):149-158.
Lakhani, K. H. and B. N. K. Davis, 1982. Multiple regression models of the distribution of Helianthemum chamaecistus in relation to aspect and slope at Barnack, England. Journal of Applied Ecology 19(2):621-629.
Lieffers, V. J. and P. A. Larkin-Lieffers, 1987. Slope, aspect and slope position as factors controlling grassland communities in the coulees of the Oldman River, Alberta. Canad. Jour. Botany 65(7):1371-1378.
Lin, T., P. M. Rich, D. A. Heisler and F. J. Barnes, 1992. Influences of canopy geometry on near-ground solar radiation and water balances of pinyon-juniper and ponderosa pine woodlands. Amer. Soc. for Photogr. and Rem. Sensing Technical Papers, Albuquerque, NM. pp. 285-294.
Lüning, K. 1971. Seasonal growth of Laminaria hyperborea under recorded underwater light conditions near Helgoland. Fourth European Mar. Biol. Symp. Cambridge Univ. Press (Cambridge, England). pp. 347-361.
Lüning, K., 1981a. Light. In Lobban, C. S. and M. J. Wynne (ed), The Biology of Seaweeds. Botanical Monographs, Volume 17. University of California Press (Los Angeles). pp. 326-355.
Lüning, K. 1981b. Photobiology of Seaweeds: ecophysiological aspects. In Proc. 10th Intl. Seaweed Symposium. Walter de Gruyter & Co. (New York). pp. 35-55.
Lüning, K. and M. J. Dring 1979. Continuous underwater light measurements near Helgoland (North Sea) and its significance for characteristic light limits in the sublittoral region. Helgoländer wissenschaftliche Meeresuntersuchungen 32:403- 424.
Lüning, K. and P. Kadel 1993. Daylength range for circannual rhythmicity in Pterygophora californica (Alariaceae, Phaeophyta) and synchronization of seasonal growth by daylength cycles in several other brown algae. Phycologia 32(5):379-387.
Lüning, K. and M. Neushul 1978. Light and temperature demands for growth and reproduction of Laminarian gametophytes in southern and central California. Marine Biology 45:297-309.
Matta, J. L. and D. J. Chapman, 1991. Photosynthetic responses and daily carbon balance of Colpomenia peregrina: seasonal variations and differences between intertidal and subtidal populations. Marine Biology 108(2):303-313.
McCree, K. J., 1972. Test of current definitions of photosynthetically active radiation against leaf photosynthesis data. Agric. Meteorol. 10:443-453.
Mel, M., 1977. Aerial Remote Sensing of Subtidal Vegetation in Southern California. Master's Thesis, University of California at Los Angeles.
Miklos, L.,I. Matecny and M. Kozova, 1991. Interpretation of the morphometric relief indices for spatial differentiation of microclimatic conditions. Ekologia (CSFR) 10(2):223-46.
Murphy, D. D. and S. B. Weiss, 1988. A long-term monitoring plan for a threatened butterfly. Conservation Biology 2(4):367-374.
Neushul, M., 1971. Submarine illumination in Macrocystis beds. In W. J. North (ed.), The Biology of Giant Kelp Beds (Macrocystis) in California. Beihefte zur Nova Hedwigia 32: 241-254.
Neushul, M., M. S. Foster, D. A. Coon, J. W. Woessner and B. W. W. Harger, 1976. An in situ study of recruitment, growth and survival of subtidal marine algae: techniques and preliminary results. J. Phycol. 12:397-408.
Neushul Mariculture, Inc., 1981. Historical Review of Kelp Beds in the Southern California Bight. Southern California Edison Company Research Report Series No. 81-RD-98. Neushul Mariculture, Inc. (Goleta, CA). 74 pp.
Nikolov, N. T. and K. F. Zeller, 1992. A solar radiation algorithm for ecosystem dynamic models. Ecological Modelling 61(3-4):149-168.
Nisbet, R. M. and J. R. Bence, 1989. Alternative dynamic regimes for canopy-forming kelp: a variant on density-vague population regulation. The American Naturalist 134(3):377-408.
North, W. J. (ed.), 1971. The biology of giant kelp beds (Macrocystis) in California. Beihefte zur Nova Hedwigia 32. 600 pp.
North, W. J. 1991. The Kelp Beds of San Diego and Orange Counties. Published by Wheeler J. North, March 15, 1991. 270 pp.
North, W. J., G. A. Jackson and S. L. Manley, 1986. Macrocystis and its environment, knowns and unknowns. Aquatic Botany 26:9-26.
Norton, T. A., 1978. Mapping species distributions as a tool in marine ecology. Proc. Royal Soc. Edinburgh 76B:201-213.
Nunez, M., 1980. The calculation of solar and net radiation in mountainous terrain. Jour. of Biogeography 7:173-186.
Okanoue, M. 1957. On the intensity of the solar radiation on any slope. J. Japan. Forest. 433-437.
Okanoue, M. and M. Makijama 1958. Computation of amount of sunshine received on a slope. J. Japan. Forest. 40-41.
Pearcy, R. W., 1983. The light environment and growth of C3 and C4 tree species in the understory of a Hawaiian forest. Oecologia 58:19-25.
Pearse, J. S. and A. H. Hines, 1979. Expansion of a central California kelp forest following the mass mortality of sea urchins. Mar. Biology 51:83-91.
Pennock, J. R., 1985. Chlorophyll distributions in the Delaware Estuary: regulation by light limitation. Estuarine, Coastal and Shelf Science 21(5):711-725.
Ramus, J. 1983. A physiological test of the theory of complementary chromatic adaptation. II. Brown, green and red seaweeds. Journal of Phycology 19:173-178.
Reed, D. C. and M. S. Foster, 1984. The effects of canopy shading on algal recruitment and growth in a giant kelp forest. Ecology 65(3):937-948.
Revfiem, K. J. A. 1978. A simple procedure for estimating global daily radiation on any surface. J. Appl. Meteor. 17:1126-1131.
Revfiem, K. J. A. 1982. Simplified relationships for estimating solar radiation incident on any flat surface. Solar Energy 28:509-517.
Rich, P. M., G. S. Hughes and F. J. Barnes 1993. Using GIS to reconstruct canopy architecture and model ecological processes in pinyon-juniper woodlands. Proceedings of the Thirteenth Annual Esri User Conference.
Robinson, N. (ed.) 1966. Solar Radiation. Elsevier (Amsterdam). 347 pp.
Rosenthal, R. J., Clarke, W. D. and P. K. Dayton 1974. Ecology and natural history of a stand of giant kelp, Macrocystis pyrifera, off Del Mar, California. Fish. Bull. 72:670-684.
Saving, S. C., P. M. Rich, J. T. Smiley and S. B. Weiss 1993. GIS-based microclimate models for assessment of habitat quality in natural reserves. Proceedings ASPRS 1993 Annual Conference, New Orleans, 3:319-330.
Seapy, R. R. and M. M. Littler, 1993. Rocky intertidal community structure on Santa Barbara Island and the effects of wave surge on vertical zonation. In F. G. Hochberg (ed.), Proc. of the Third California Islands Symposium: Recent Advances in Research on the California Islands. Santa Barbara Museum of Natural History (Santa Barbara, CA). pp. 273-292.
Sieburth, J. M. and A. Jensen 1968. Studies on algal substances in the sea. I. Gelbstoff (humic material) in terrestrial and marine waters. J. Exp. Mar. Biol. Ecol. 2:174-189.
Sjoberg, R. W. and B. K. P. Horn, 1983. Atmospheric effects in satellite imaging of mountainous terrain. Applied Optics 22(11):1702-1716.
Smith, R. C., W. Wheeler and L. Srivastava, 1983. Seasonal photosynthetic performance of Macrocystis integrifolia (Phaeophyceae). Jour. Phycol. 19:352-359.
Smith, R. C., et al., 1987. Optical characterization of primary productivity across a coastal front. Mar. Biology 96:575-91.
Statistical Sciences 1993. S-Plus for Windows User's Manual, Volumes 1 and 2; S-Plus for Windows Version 3.2 Supplement; S-Plus Programmer's Manual, Version 3.1. Statistical Sciences Inc. (Seattle WA).
Steinman, A. D., P. J. Mulholland, A. V. Palumbo, T. F. Flum and D. L. DeAngelis, 1991. Resilience of lotic ecosystems to a light-elimination disturbance. Ecology 72(4):1299-1313.
Taylor, A. H. and J. A. Stephens, 1983. Seasonal and year-to-year changes in the temperatures of the English Channel and the Southern North Sea, 1961-1976: a budget. Oceanologica Acta 6(1):63-72.
Temps, R. C. and K. L. Coulson, 1977. Solar radiation incident upon slopes of different orientations. Solar Energy 19:179-184.
Thom, R. M. and R. G. Albright, 1990. Dynamics of benthic vegetation standing-stock, irradiance and water properties in central Puget Sound. Marine Biology 104(1):129-141.
Tyler, J. E. and R. C. Smith, 1970. Measurements of Spectral Irradiance Underwater. Gordon and Breach Sci. Publishers (New York, NY). 103 pp.
Walsh, J., D. Dieterle and W. Esaias 1987. Satellite detection of phytoplankton export from the mid-Atlantic Bight during the 1979 spring bloom. Deep Sea Research 34(56):675-703.
Weiss, S. B., D. D. Murphy and R. R. White, 1988. Sun, slope and butterflies: topographic determinants of habitat quality for Euphydryas editha. Ecology 69(5):1486-1496.
Weiss, S. B., D. D. Murphy, R. R. White and A. D. Weiss 1993. Estimation of population size and distribution of a threatened butterfly: GIS applications to stratified sampling. Proceedings of the Thirteenth Annual Esri User Conference.
Wheeler, W. N., 1978. Ecophysiological Studies on the Giant Kelp, Macrocystis. Ph D. Dissertation. Biology Department. University of California at Santa Barbara. 179 pp.
Wilkinson, L., M. Hill, J. P. Welna and G. K. Birkenbeuel, 1992. Systat, the System for Statistics. Systat, Inc. (Evanston, IL). 750 pp.
Wing, S. R., J. J. Leichter and M. W. Denny, 1993. A dynamic model for wave-induced light fluctuations in a kelp forest. Limnol. Oceanogr. 38(2):396-407.
Woods, J. D. and R. Onken, 1982. Diurnal variation and primary production in the ocean- preliminary results of a Lagrangian ensemble model. Jour. Plankton Res. 4(3):735-756.
Yeaton, R. I. and M. L. Cody, 1979. The distribution of cacti along environmental gradients in the Sonoran and Mohave deserts. Jour. Ecol. 67:529-541.
Youtie, B. A., B. Griffith and J. M. Peek, 1988. Successional patterns in bitterbush habitat types in north-central Washington. Jour. of Range Management 41(2):122-126.
Zimmerman, R. C. and D. L. Robertson, 1985. Effects of El Niño on local hydrography and growth of the giant kelp, Macrocystis pyrifera, at Santa Catalina Island, California. Limnol. Oceanogr. 30(6):1298-1302.
William W. Bushing (bushing@lifesci.ucsb.edu)
Ecology, Evolution and Marine Biology
University of California
Santa Barbara, CA 93106
and
Director of Science, Education and Ecological Restoration (SEER)
Catalina Island Conservancy
P. O. Box 2739
Avalon, CA 90704
(310) 510-2595 ext. 134
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}