William W. Bushing
This study utilized a GIS to investigate the influence of submarine and terrestrial topography on the distribution of giant bladder kelp around an island off the southern California coast. Macrocystis pyrifera is an important species in the marine environment, serving both as structural habitat and food for a wide range of marine invertebrates and fishes. Santa Catalina Island's highly dissected, 54-mile leeward and windward coastline offers a wide range of microhabitats with respect to storm exposure, temperature and the nearshore light regime. GIS data layers included a digital terrain model, digital bathymetric model, submarine slope, submarine aspect, bottom relief, a series of solar insolation models (monthly, seasonal, annual), significant wave height, and kelp distribution maps from several survey years. Definite patterns in distribution were observed relative to aspect, depth, slope, temperature, surface insolation and bottom insolation. GIS overlay methods used on the multi-temporal kelp distribution maps generated a layer representing the spatial persistence of kelp. Correlations between persistence and the physical variables were used to characterize regions suitable for designation as marine reserves. The most interesting "finding" in this study was that many marine scientists are not yet convinced GIS can offer them a new tool to investigate ecological phenomena at different spatial and temporal scales from those conventionally employed in field or laboratory research.
Giant kelp (Macrocystis pyrifera) is a species of marine alga found along the Pacific coast of North America from central California to Baja California (Abbott and Hollenberg 1976). Although it begins life as a microscopic spore at the ocean floor, this species may grow to lengths of 60 m with its upper fronds forming a dense canopy at the surface. Giant kelp prefers depths less than 40 m, temperatures less than 20 C, hard substrate, and bottom light intensities about 1% that of the surface (North 1971, Foster and Schiel 1985). Macrocystis plays an important role in the marine environment by providing food and habitat for a wide range of marine invertebrates and fishes in southern California. Forests of giant kelp may support millions of individual organisms and more than 1,000 species of marine plants and animals (North 1971, Earle 1980, Foster and Schiel 1985).
Santa Catalina Island is one of the eight Channel Islands located off the southern California coast (figure 1). The island is oriented in a NW-to-SE direction relative to the prevailing westerly and northwesterly swell and wind (figure 2). This orientation results in a high energy exposed coastline on the southwest-facing windward side and a lower energy, more protected environment on the northeast-facing leeward side. In addition, the island's 54-mile coastline is highly dissected with small bays, points and other coastal features. Combined, these geographic characteristics result in a wide range of microhabitats with respect to storm exposure, temperature, light and other environmental parameters.

Most of southern California's giant kelp is found along the coasts of the offshore Channel Islands, including Catalina. Anecdotal and personal observations suggest reductions in the distribution of Macrocystis around this island over the past 25 years. Similar trends have been noted elsewhere in the region, and have been attributed to sewage pollution, sea otter hunting, sea urchin grazing, elevated temperatures associated with El Nino events, and other causes (Hodder and Mel 1978; Neushul Mariculture Inc. 1981; Harger 1983; North 1991).
The loss of kelp, and the important marine habitat that it represents, has been one factor stimulating interest in the establishment of marine reserves around Catalina Island. Several existing reserves have already been designated based largely on recreational or educational access. These include Lover's Cove and Casino Point designated by the City of Avalon, Toyon Bay (Catalina Island Marine Institute) and a reserve at Blue Cavern Point associated with USC's Wrigley Marine Science Center at Big Fisherman Cove (figure 3). The California State Water Resources Control Board has also designated stretches of the island's coastline as Areas of Special Biological Significance (ASBS) affording some additional protection (SWRCB 1979, 1981).
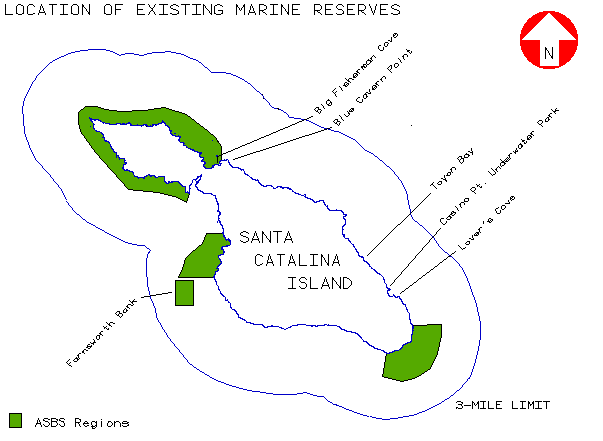
With the exception of the Farnsworth Bank ASBS, these reserves are all located on the leeward side of the island near existing population centers. Island-wide studies using a geographic information system (GIS) have indicated that most of the island's kelp beds are located on the more exposed windward coast (Bushing 1994). This suggests that selection of future marine reserves should be done on the basis of ecological criteria such as disturbance, persistence and metapopulation characteristics, rather than strictly on issues of access or recreational value.
This study utilizes the same GIS with additional data layers to elaborate on the preliminary research referenced above. The new data layers include models of wave and storm exposure, solar insolation and the persistence of kelp around the island. These layers allow a better understanding of ecological disturbance factors and the ability of kelp to recover from them. The persistence layer incorporates multi-temporal kelp maps allowing the analysis of kelp distribution over time relative to the other variables.
Ecologically, the term disturbance has been defined as the discrete removal of individual organisms from a biological population, a detectable change in population response to sudden changes in resources, or an event causing a community or ecosystem characteristic (eg, species diversity) to diverge from its normal range (Forman and Godron 1983, Bazzaz 1983, Wallace 1990, Gore et al. 1990). Removal of kelp plants by storms is an example of the first, reductions in growth or biomass due to elevated El Nino temperatures the second, and decreases in species diversity of the marine invertebrates associated with kelp the third. The "disturbance regime" includes the type, areal extent, magnitude, frequency, predictability and turnover rate of disturbance events through time within a given landscape (Forman and Godron 1983, Sousa 1984). The disturbance regime is relative to the characteristics of both the local landscape, and the species of interest. Environmental factors influencing the disturbance of Macrocystis may be biological or physical in nature.
Biological factors include grazing by invertebrates or fish, competition from other algae, encrusting growth and diseases (North 1971, Foster and Schiel 1985). Marine invertebrates known as sea urchins are often cited as agents of disturbance in kelp bed systems globally (eg, Mann and Breen 1972; Lawrence 1975, Hagen 1983) and in southern California (eg, Leighton et al. 1966, North and Pearse 1970, Tegner 1980). Several species of sea urchins are known from Catalina waters. Based on a model by Harrold and Reed (1985), and mechanisms discussed by Ambrose et al. (1993), conditions around Catalina are generally favorable for sea urchin overgrazing due to low nutrients, warm temperatures and exposure to storms on the windward side. However, events similar to those observed on the other Channel Islands (Richards et al. 1993, Ambrose et al. 1993) are rarely observed in our waters based on published reports (Engle 1994, McAlary and McFarland 1994) and personal observations. Macrocystis pyrifera exhibits such strong competitive advantage over other local algae that competition is rarely a problem (Dayton et al. 1992).
The absence of significant local biotic disturbance makes physical factors of primary importance (North 1968, 1971; Foster and Schiel 1985). Warm water episodes such as El Nino also have serious effects on the growth and survival of giant kelp (eg, Dayton and Tegner 1984, Gerard 1984). These large-scale events play a role in the disturbance regime directly through the physiological stress associated with elevated temperature or indirectly due to associated phenomena (eg, reduced nutrients or strong storms). On a local scale, exposure to wind, waves, swell and storm events plays an important role in Catalina's disturbance regime. Storms impact the island's windward side primarily from the NW or W during winter, but may also strike from the south during summer or on the leeward side during Santa Ana winds.
A species' response to such disturbance events determines its ability to persist locally through time. Ecological persistence may be simply defined as the absence of local extinction (Rabinowitz et al. 1989) or the continued existence of a species over time (Connell and Sousa 1983). Two components of a species' stability are resistance, the ability to withstand disturbance, and resilience, the ability to return to its pre-disturbance state (Connell and Sousa 1983). The spatial heterogeneity of the physical habitat may play a significant role through the presence of microhabitats serving as refugia (Sousa 1984), high complexity (Pearsons and Li 1992), high nutrient supplies (Steinman et al. 1991) and other factors.
This study utilizes a GIS to focus on environmental factors that appear to confer higher persistence for kelp around Catalina. The physical disturbance regime is characterized based on available data and models. Multi-temporal kelp distributions are used to generate levels of persistence. The distribution of these levels is evaluated statistically to determine which factors contribute to kelp's stability geographically. It is hypothesized that in regions of higher disturbance kelp will favor sites in protected coastal aspects, deeper water, shallower slopes and higher bottom relief and show higher persistence there. Finally, this information will serve to generate guidelines for identifying kelp beds suitable for designation as future marine reserves.
GIS data layers required to address the issues in this study include several kelp bed distribution maps, bathymetry (depth), submarine aspect, submarine slope, bottom substrate relief, solar insolation and significant wave height. Data layers were all registered to base maps of the island's shoreline manually digitized from the four USGS 7.5' quad maps covering Santa Catalina Island at a scale of 1:24,000 using UTM-11 coordinates. Topographic contours were manually digitized from the same maps at 50-250' intervals depending on local topography. Bathymetric contours to a depth of 100 fathoms were digitized from NOAA chart 18757 ("Santa Catalina Island"). A digital terrain model was interpolated (IDW) from the combined topographic and bathymetric contours at 20 m resolution to coincide with the spatial resolution of the SPOT satellite imagery used. Aspect (figure 4) and slope data layers were generated from the resulting image.

The initial kelp bed distribution map, dated about 1980, was provided by Southern California Edison in ArcInfo EXPORT format (figure 5). Three additional distribution layers were generated from: (a) vector point data for 1934 from NOAA's National Ocean Survey database; (b) polygons manually digitized from a 1977-78 kelp distribution map (SWRCB 1979); and (c) a raster-to-vector conversion of kelp presence from a 1990 SPOT satellite digital image in the near infrared (NIR). Polygon buffers were created at a distance of 60 m from each point in the 1934 data using BUFFER. Kelp's high reflectance and waters strong absorption in the NIR allowed the delineation of kelp canopy in the satellite image using image processing techniques to create a distribution map.
All four distribution maps were converted to binary raster data layers indicating presence (1) or absence (0) of kelp in each using POLYGRID. The four layers were composited using GRID overlay methods so each cell in the resulting "persistence" layer represented the number of survey years in which kelp was found at that geographic position. The resulting values ranged from 0 (kelp present in no survey years) to 4 (kelp present in all four survey years). A similar method has been used by Southern California Edison to look at the persistence of kelp near its San Onofre generating plant (Albert Lin, pers. comm.).
Because bottom relief is a potentially important factor for kelp distribution, GIS layers representing indices of relief were generated. The aspect layer was re-classed into twenty-four 15-degree categories (eg, 0-14 degrees). The resulting image was imported into ERDAS image processing software and rectangular "diversity" or "texture" (Star and Estes 1990) filters were applied at several spatial resolutions to model the heterogeneity in the region of each raster cell. The variable dimensions of the spatial filters permitted the investigation of bottom relief at three separate spatial scales. The 3x3, 5x5 and 10x10 pixel filters yielded an index of bottom relief at spatial scales of about 30-40, 50-70 and 100-140 m respectively from the subject cell.
Texture analysis, the application of spatial filters to ecological landscapes, has become popular with the emergence of GIS and image processing as tools for the biologist. It offers one means of quantifying landscape characteristics for subsequent analysis. These techniques have been used to calculate measures of landscape heterogeneity, habitat diversity and habitat fragmentation (eg, Weaver and Kellman 1981, Ripple et al. 1991)
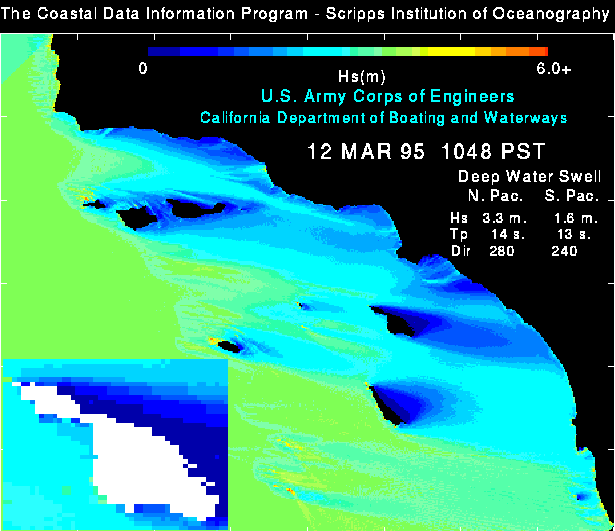
Storm exposure was incorporated into the GIS from a model developed by O'Reilly and Guza (1993) employing submarine bathymetry and swell characteristics to estimate significant wave heights (Hs) in the Bight. Hs is defined as the mean height of the largest 1/3rd of observed waves (Pond and Pickard 1983). An image modelling Hs at 200-m resolution for a peak storm event on March 12, 1995 (1048 PST) was re-created by William O'Reilly and provided for this analysis (figure 6). The image was then printed in color at a scale that permitted digitization of the boundaries between Hs values into vector polygons encoded with the corresponding Hs value. These polygons were registered to the base layers and incorporated into the GIS. The polygons were also converted to raster data for cell-by-cell analysis.
Because kelp canopy responds to wave energy as well as wave height, Hs was also quantized into three wave energy levels based on the following equation from Pond and Pickard (1983):
E = (rho * g * Hs^2)/8
Since wave energy is proportional to the square of Hs, values of Hs < 1.40 were assigned to energy level 1, 1.40 < Hs < 2.00 to level 2, and Hs > 2.0 to level 3. These classes permitted statistical analysis of differences in slope and depth as a function of Hs and wave energy.
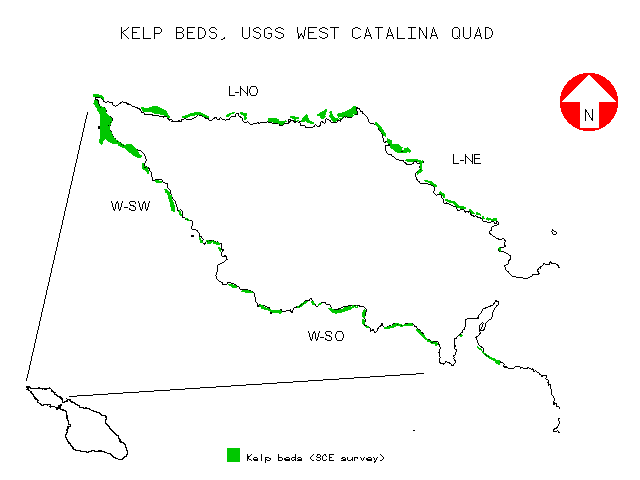
Many of the data layers were stored both as polygon and raster data to facilitate the analyses in this study at differing spatial scales. The analyses here focus only on the West Catalina USGS quad. The shoreline for this quad was classified into two primary geographic regions (West windward and West leeward), as well as four subregions (W-SW and W-SO on the windward side and L-NO and L-NE on the leeward side) based on the prevailing direction each coast faced.
Although analysis was conducted using the vector polygon layers, much of the information presented here looked at the data on a cell-by-cell basis due to the influence of smaller-scale (~20-m) heterogeneity on kelp distribution. Binary raster masks representing kelp distribution from the 1980 SCE survey were created at 20-m resolution from the original polygon data for both leeward and windward coasts. The 20-fathom bathymetric contour was chosen to define the maximum expected depth limit for kelp. The region between this contour and the shoreline represents the potential area that might be occupied by kelp. For this study it is referred to as the "photic shelf" since its lower limit is generally considered to be light-limited. A second set of binary masks corresponding to the "photic shelf" on each coast was created using the shoreline and 20-fathom contours.
Coincident data was extracted from the raster GIS layers for aspect, depth, slope, bottom relief, and significant wave height using the kelp and photic shelf binary masks and GRID overlay techniques. This data was analyzed within the GIS and the resulting grids and binary masks were converted to ASCII format for import into Foxpro databases representing the windward and leeward sides. Each data layer (eg, binary kelp distribution mask, extracted aspect) was imported as a separate field in the database so each record represented a single geographic location incorporating the associated data in multiple fields. Several programs were written in the xBASE computer language to further analyze the data on a cell-by-cell basis.
Two important statistics were evaluated for each data layer: kelp frequency and kelp percent cover. In this paper the phrase "kelp frequency" refers to the percentage of total kelp along each coastline with a given data level (eg, 5% of all kelp on the West leeward coast was found at a depth of 18 m). The phrase "percent cover" refers to the percent of the corresponding "photic shelf" covered by kelp for a given value of an environmental variable (eg, 20% of all 8 m depths on the West windward coast were covered by kelp).
Statistical tests were run on the extracted data to determine which environmental factors appear to be related to the persistence of kelp on the windward and leeward coasts. The relational databases for each coast were imported directly into Systat statistical software. Tests for statistically significant differences were then run for the different persistence levels.
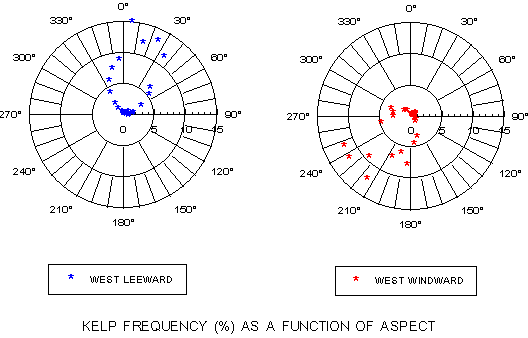
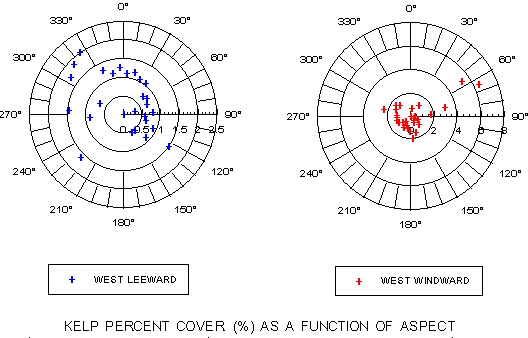
Kelp frequency and kelp percent cover as a function of submarine aspect are presented graphically for the West Catalina leeward and windward coastlines. Due to the prevailing NW-to-SE trend of the island, the frequency distribution (figure 7) shows strong peaks at 0-40 degrees on the north- and northeast-facing leeward side and 210-250 degrees on the south- to southwest-facing windward side. When distribution relative to aspect is analyzed in terms of percent cover, a different picture emerges (figure 8). Marked peaks are observed at ~120 and 300-330 degrees on the leeward side and 50-90 degrees on the windward side.
Depth data was also presented for the leeward and windward coasts. Kelp frequency distribution (figure 9) on the leeward side peaked at statistically significant (p < 0.001) deeper depths than on the windward side. Mean and maximum depth values were 11.6 and 41.8 m on the leeward side versus 7.1 and 24.1 m on the windward side. Percent cover (figure 9) also indicated the same pattern with maximum values at deeper depths on the leeward side (8-9 m) then the windward (3-5 m).

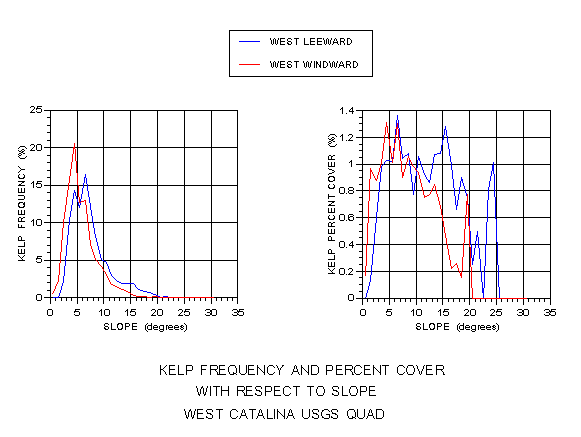
Figure 10 depicts the results for kelp frequency and percent cover relative to submarine slope. Kelp frequency also shows statistically significant differences (p < 0.001) with kelp found on shallower slopes on the windward side than the leeward. Mean values were 2.9 degrees on the windward side and 3.6 on the leeward. Percent cover shows generally higher values at lower slopes on the both coasts, but an avoidance of steep slopes on the windward side.
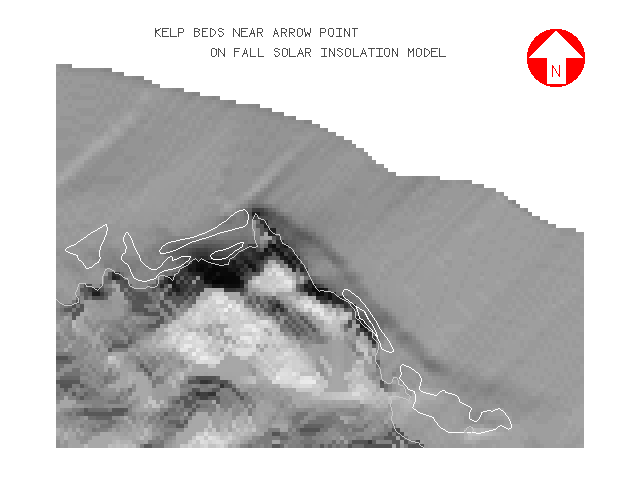
The persistence layer is illustrated by figures 11 and 12 showing the regions of Arrow Point on the leeward side, and the extreme West End of the island. It can be seen from the legend that very few cells had kelp present in all four survey years. Mean persistence on the exposed windward side was higher at 2.38 compared to 1.83 for the more protected leeward side. On the windward side 78% of all kelp was found in levels 2 and 3, while on the leeward side 85% was in levels 1 and 2.
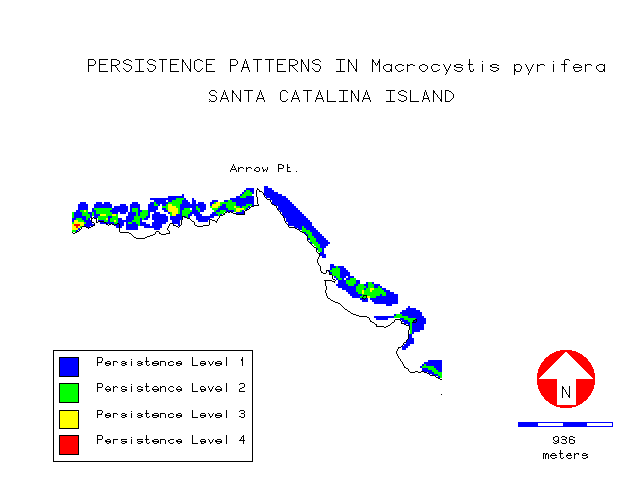
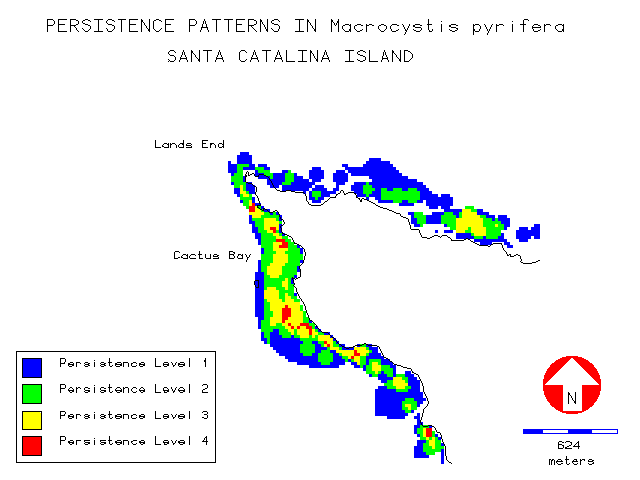
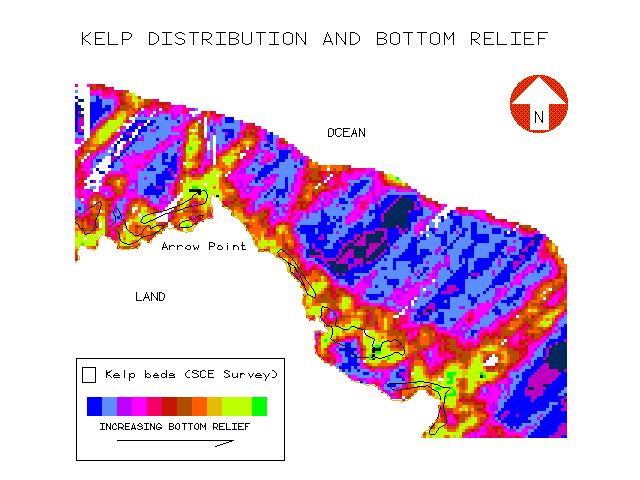
Visually, kelp appeared to have a strong affinity for regions of increased bottom relief such as offshore reefs and areas around coastal points and headlands as illustrated in the region near Arrow Point (figure 13):
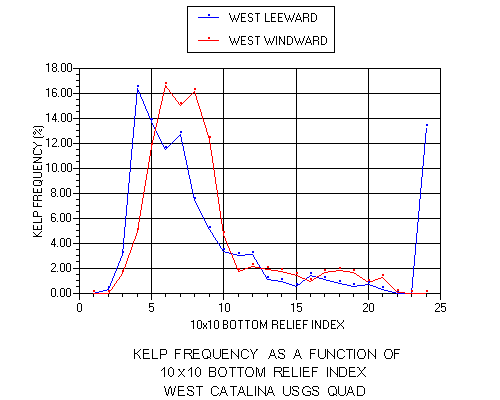
Although the mean values relative to bottom relief were lower on the windward side, kelp frequency peaked at higher values (6-8) there than on the leeward side (4-7). The mean values were significantly higher for kelp than for the corresponding shelf on both sides, but test statistics indicate the relationship was stronger for the windward side (p l< 0.001) than the leeward side (p < 0.010). There were also detectable trends in average bottom relief relative to aspect as presented in Figure 14.
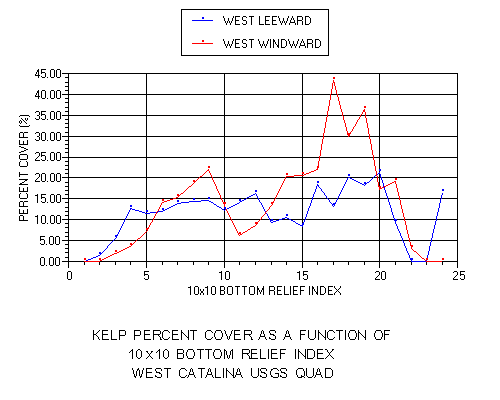
The presentation of kelp percent cover data is more revealing (figure 15). The leeward side exhibits a more uniform distribution (10-20%) relative to bottom relief. No prominent peaks are noted, although there is a slight trend for increasing percent cover with increasing bottom relief. On the windward side percent cover was low (0-10%) in regions of low bottom relief (1-5), quite variable (7-22%) at moderate relief (6-16) and exhibited strong peaks (30-45%) in regions of high relief (17-19). Similar trends were noted at all three spatial scales.
A casual glance at the significant wave height (Hs) image for March 12, 1995 (figure 6), confirms the expected difference between the exposed windward and protected leeward coasts. Closer inspection also reveals the development of strong "swell shadowing" on the southern portions of the windward coast, and to a lesser extent along that portion of the West windward coast that faces almost directly south (W-SO).
The frequency distribution for kelp on the windward and leeward sides relative to Hs is consistent with these general patterns (figure 16). About 90% of kelp on the leeward side was found at lower values of Hs (0.24-1.41 m). On the windward side, distribution fell into two ranges (1.41-1.70 and 2.21-2.62 m), reflecting the south and southwest exposures along this coastline, with nearly 70% of the kelp in the upper range.
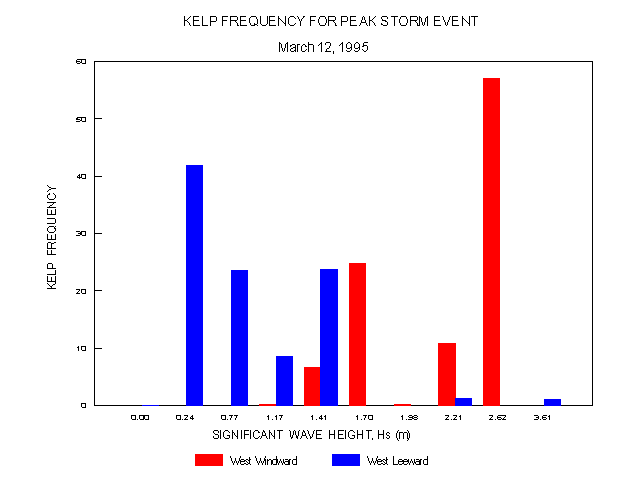
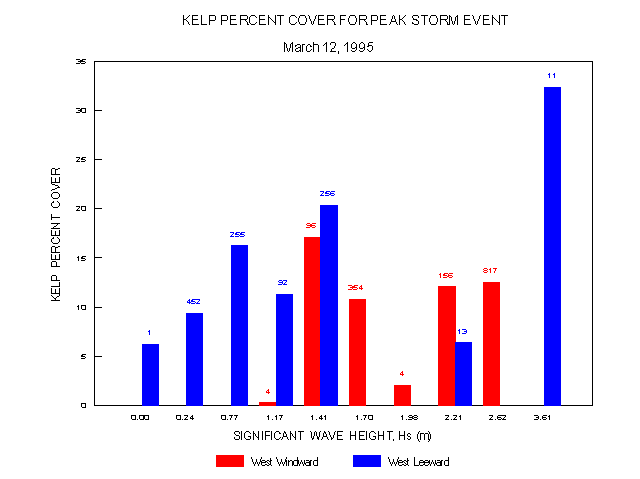
Percent cover of kelp (figure 17) showed some significant divergence from the frequency distribution. On the leeward side, values were higher (11-32%) at moderate (0.77-1.41) and high (3.61 m) values of Hs while on the windward side the highest percent cover (17%) was near the lower Hs range (1.41 m).
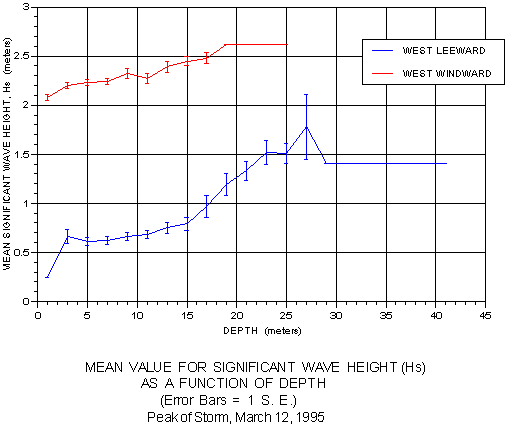
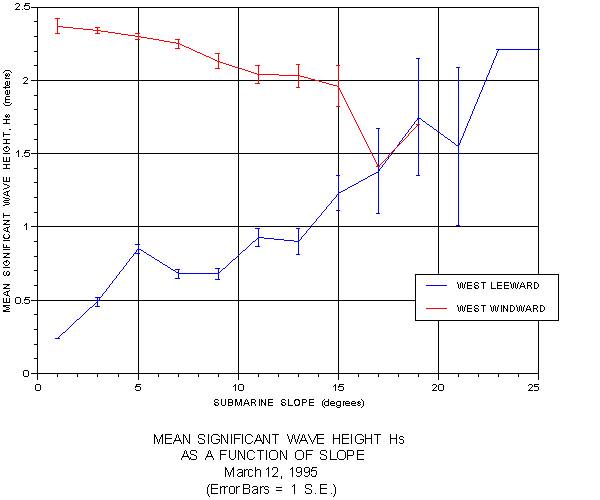
Very revealing patterns emerged when the mean value for Hs was graphed as a function of depth (figure 18) and slope (figure 19). In the first graph, mean Hs steadily increases on both the windward and leeward sides with increasing depth. Plotted against slope, mean Hs steadily declined on the windward side as slope increased, but the reverse trend was noted on the windward side. Statistical tests were applied to the frequency distributions for depth and slope at the three levels of swell energy. Significant differences (p < 0.001) existed between all three swell energy classes for both depth and slope on the protected leeward side, but only between energy classes 2 and 3 on the exposed windward side.
Analysis of persistence as a function of storm exposure, represented by wave energy (Hs^2), generated results consistent with expectations. Persistence exhibited a statistically significant difference for each of the three wave energy levels (0.00-1.40; 1.41-1.99 and 2.00-4.00 m) on the exposed windward side, but none on the leeward side. The following table presents this data in the form of pairwise comparison probabilities:
Table 1: Pairwise comparison matrix of statistical significance in differences between kelp persistence as a function of swell energy on the West windward and leeward coasts (energy class levels as defined in the text).
Windward Side Leeward Side Energy Class 1 2 3 1 2 3 1 1.000 1.000 2 0.005 1.000 0.081 1.000 3 0.002 0.020 1.000 0.881 0.964 1.000
Looking at depth as a factor in persistence, there appears to be little significant influence on either side of the island. On the windward side the only highly significant difference was noted between persistence level 1 and levels 2-4, although the difference between levels 2 and 3 approached p = 0.05. On the leeward side none of the pairwise comparisons in the probability matrix showed any significant difference.
Table 2: Pairwise comparison matrix of statistical significance in differences between kelp depth distributions as a function of persistence level on the West windward and leeward coasts.
Windward Side Leeward Side Persistence 1 2 3 4 1 2 3 4 1 1.000 1.000 2 0.000 1.000 1.000 1.000 3 0.000 0.059 1.000 0.995 0.986 1.000 4 0.000 0.999 0.362 1.000 0.999 0.999 0.997 1.000
Slope appears to play a more important role as a factor in determining persistence. Based on the calculated probabilities, differences in slope have a high statistical significance between all four levels of persistence on the windward side. On the leeward side, persistence levels 1-2 and 2-3 show significant differences while 3-4 do not. However, much of the observed difference is attributed to the slope characteristics of level 2. The following matrix presents these probabilities:
Table 3: Pairwise comparison matrix of statistical significance in differences between kelp slope distributions as a function of persistence level on the West windward and leeward coasts.
Windward Side Leeward Side Persistence 1 2 3 4 1 2 3 4 1 1.000 1.000 2 0.000 1.000 0.000 1.000 3 0.000 0.000 1.000 0.994 0.000 1.000 4 0.000 0.000 0.000 1.000 0.999 0.842 0.997 1.000
The last topographic variable investigated relative to persistence was the 10x10 bottom relief index. The patterns follow the similar trends to those for slope and swell. Kelp distributions on the windward side show statistically significant differences between levels 1-2 and 2-3, but not between 3-4 for bottom relief. The leeward side pairwise probability matrix shows no significant difference between any of the persistence levels.
Table 4: Pairwise comparison matrix of statistical significance in differences between kelp bottom relief distribution as a function of persistence level on the West windward and leeward coasts.
Windward Side Leeward Side Persistence 1 2 3 4 1 2 3 4 1 1.000 1.000 2 0.000 1.000 1.000 1.000 3 0.012 0.024 1.000 0.928 0.884 1.000 4 0.985 0.000 0.126 1.000 0.742 0.728 0.837 1.000
The response of Macrocystis pyrifera to the disturbance regime around Santa Catalina Island, and therefore its persistence in the face of disturbance, is a complex issue due to the varying geographic impact of disturbance events and the wide range of microhabitats afforded by the island's coastline. The spatial heterogeneity of the physical environment appears to be a major factor in determining where giant kelp survives. Coupled with this is the seasonally and geographically variable nature of disturbance events on the island.
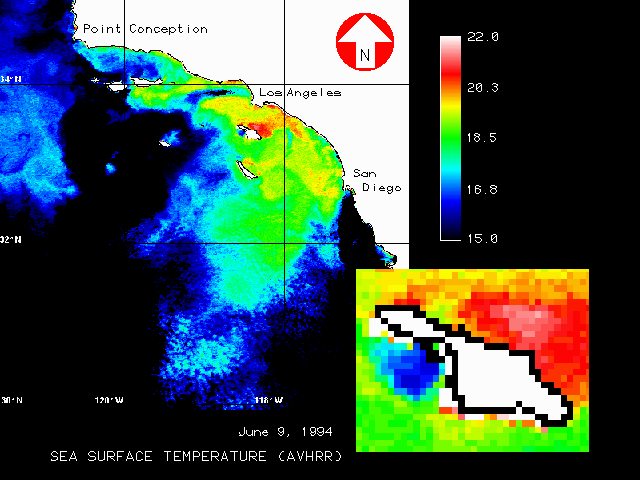
The effects of El Nino events on Macrocystis have established the influence of temperature as a major disturbance factor for Macrocystis over large spatial scales. The waters surrounding Catalina are the warmest in the Channel Islands region, and temperatures regularly exceed 20 C during the warmer months. Thus kelp around the island is near the limits of its preferred temperature range. In another study (Bushing 1995), sea surface temperature (SST) data from AVHRR satellite imagery archived by NOAA (CoastWatch 1995) revealed patterns in the local distribution of ocean temperature of potential significance for Macrocystis. Temporal and spatial variability (both regional and local) in temperature was highest during the summer months, while winter values were more uniform. Average summer temperatures on the leeward side were uniformly and significantly warmer than on the leeward side. Warm water bodies were observed frequently on the leeward side, and cooler water regions were apparent on the windward side in many images (figure 20).
The observation that significant temperature differences exist between the windward and leeward sides of the island suggests possible implications relative to the recovery of kelp from El Nino disturbances. Reductions in kelp canopy noted for the leeward side began about 1976 coinciding with a mild El Nino. Ocean temperatures since that time have remained above normal (Norton et al. 1985, Stevens 1994). The consistently higher temperatures on the island's leeward side, and the resultant physiological stress, may be a factor in explaining why kelp has not recovered there, yet appears to be doing well on the windward side.
However, at smaller scales surge and waves associated with storms may be the most important cause of adult mortality in Macrocystis, and therefore a prime disturbance factor. The windward side of Santa Catalina Island represents an exposed outer coastline receiving the incoming storm energy with little protection. Its steep submarine slopes compared to those of the mainland may accentuate the impact of incoming storms. For a given wavelength, wave energy is released as a function of depth, being stronger in shallower water. Along steep coasts, wave energy may be dissipated closer to shore where kelp usually grows, rather than farther away as along the more gently-sloping mainland shelf. Regions of higher bottom relief may offer some refuge by breaking up the incoming swell. Therefore the location relative to storm direction (aspect), depth, the slope and relief of the ocean floor may all play a role in determining the ability of a kelp bed to resist storm disturbance.
Many authors have stressed the importance of coastal aspect in determining the size or viability of kelp beds (Hodder and Mel 1978, Harger 1983, Reed and Foster 1984, North 1991). The analysis of kelp distribution around Catalina relative to aspect appears to validate the hypothesis that kelp favors regions offering some shelter from the prevailing storm or swell direction. Anomalously high percent cover at aspects facing away from the prevailing coastal directions on both windward and leeward sides suggest kelp may have higher survivorship there. Such regions in the lee of points or headlands, offshore reefs or other coastal features may serve as refuges from storm impact.
Contrary to expectations, kelp on the exposed windward side did not favor deeper water where surge effects would be minimized. Instead, its distribution was in uniformly shallower depths than on the corresponding leeward side. This finding has been interpreted as due to increased turbidity on the windward coast due to the re-suspension of bottom sediments in the higher energy environment (Bushing 1994). North (1991) stated that the outer edges of mainland kelp beds usually lie at depths of 18 m or less except where the water may be very clear, allowing light to penetrate deeper. This also coincides with the surge zone defined by Quast (1971). Kelp on the windward side falls almost entirely within this vertical zone subject to more frequent disturbance by storm, suggesting higher mortality and turnover typical of shallower depths (Dayton and Tegner 1984, Seymour et al. 1989).
Differences in topography resulting from the uplift of the island's land mass have resulted in shallower submarine slopes on the windward side. The photic shelf there is more like that of the adjacent mainland than the island's steeper leeward side. Incoming waves and surge respond differently to the shallow and more steeply-sloping bottom topography. Energy may be dissipated more gradually on shallow slopes resulting in less destructive water motion at comparable depths. The higher observed percent cover at shallow slopes on the windward side supports the hypothesis that kelp would favor such regions in the face of storm exposure.
Geographic relief due to topography has long been known to influence the distribution of terrestrial plants (eg, Holland and Steyn 1975, Krcho 1991). Kelp beds are also linked directly to submarine topography, with areas of greater bottom relief generally supporting more kelp (SWRCB 1979, Harger 1983). In this study the more uniform distribution relative to bottom relief on the leeward side suggests substrate occupation without regard to bottom relief. On the windward side, higher percent cover in areas of moderate to high relief suggests a preference for such regions in the face of increased exposure to storms.
The incorporation of storm exposure, as measured by the significant wave height, offers an improvement over the preliminary research reported earlier (Bushing 1994). Without this element, only subjective statements could be made about the relationship between environmental variables and the potential disturbance regime. Although the storm event selected represents but one of a wide range of events due to varying wave heights and swell directions, it does allow quantitative data to be used in this analysis. The findings from this study suggest kelp on the leeward side favors regions of moderate rather than low Hs, indicating a preference for some water motion. The high percent cover at lower values of Hs on the windward side strongly suggests that regions protected from direct storm impact are favored there.
The evaluation of mean Hs as a function of depth and slope also confirms the original hypotheses regarding disturbance. Steadily increasing values of Hs with increasing depth on both leeward and windward coasts may be interpreted as a result of higher mortality in shallower waters due to storm exposure. Kelp may persist in regions of shallow water only when wave and storm exposure is low. At deeper depths, the energy of incoming swell is felt less strongly so kelp may persist there despite increased values of Hs.
The results relative to slope suggest responses to it are more regional in nature. The decreasing Hs values with increasing slope on the exposed windward side also confirm the original hypotheses. In regions of shallow slope, where wave energy may be dissipated gradually, kelp may persist in areas of higher Hs while on steeper slopes they require quieter waters. The reverse trend on the leeward side suggests that on this more protected coast other environmental factors may be more significant.
Using these same variables (aspect, depth, slope, bottom relief and storm exposure) to evaluate "persistence" first necessitates a consideration of resistance and resilience relative to the model used. Macrocystis is a perennial alga, with individual plants surviving for several years and entire kelp beds potentially even longer. Another factor is the differential ability of adult sporophytes near the surface and the microscopic stages near the bottom to survive in the face of disturbance. A storm which detaches the adult plant due to the force applied to its canopy may leave the young stages relatively undisturbed. The subadult stages at the bottom may also remain vegetative until conditions are appropriate for their growth, just as plant seeds remain in the soil bank for several years. In addition Macrocystis is a strong competitive dominant compared to most algae. Kelp beds are known to fully recover from disturbance within 14 months (Kennelly and Underwood 1993).
The "persistence" model employed in this study utilized widely-separated (1934, 1978-79, 1980 and 1990) temporal data. The time gaps in the data are too wide to determine whether the levels of "persistence" represented in this model measure the resistance of kelp, and therefore its ability to withstand disturbance, or its resilience, the ability to recover. In the future, additional temporal maps will be incorporated into the model to better evaluate this. Given these limitations, the model can be used to evaluate the following trends.
The influence of storm exposure on kelp persistence is consistent with expectations. On the higher energy windward coast, significant differences in persistence were noted for differing levels of wave energy. Locations that offer more protection may serve as refuges and exhibit higher survivorship. Kelp in such refugia may help re-populate nearby sites where more direct storm exposure has eliminated kelp beds. The absence of significant differences in persistence on the protected leeward side may simply reflect the smaller amplitude in Hs observed there. Reice et al. (1990) argued that the most frequently and predictably disturbed sites should demonstrate the highest resilience. The findings in this study are consistent with that hypothesis.
Depth did not appear to be a highly significant favor in persistence based on the pairwise probability matrix in Table 2. The restricted depth distribution on the windward side may mask any linkage with survivorship as suggested by the presence of some measurable differences. On the more protected leeward side, where storms may be less of a factor than large-scale disturbance due to elevated ocean temperatures, the value of deep water refugia may be diminished since canopy can't persist in the warmer surface waters.
The strong relationship between persistence and submarine slope (Table 3), especially on the windward side, adds further confirmation to its importance as a physical habitat factor. All persistence levels exhibited significant differences in slope on that coast. Although not as important a factor on the leeward side, slope still appears to play a role in conferring greater stability in kelp beds.
Finally, bottom relief also appears to be a determining factor in the stability of kelp on the windward side (table 4). In the face of higher wave energy, the protection afforded in regions of greater relief on the windward side may confer sufficient advantage to allow kelp to resist storms. The absence of any significant difference on the leeward side is equally informative. On this more protected coast, the absence of strong storm exposure may minimize the value of bottom relief.
As suggested in the introduction, the ability of kelp to recover from disturbance may depend on the physical heterogeneity of the environment relative to topographic factors and the presence of refugia from direct storm effects. Given comparable disturbance regime, kelp may persist in one site and not another if resources allow it to repair damaged tissue or grow at rates sufficient to keep up with losses due to disturbance (Jackson 1977). Additional resource factors which may influence this are temperature, light and nutrients. Temperature was addressed above. The latter two factors, both essential for photosynthesis, have been addressed in detail previously using the GIS (Bushing 1994, 1995), but will be summarized for consideration here.
In the earlier studies, kelp distribution was evaluated relative to the light regime using models of solar insolation (figure 21). The windward side had uniformly higher levels of surface insolation throughout the year while the light regime on the leeward side was lower due to shading of the nearshore waters by the island land mass. Kelp on the leeward coast favored sites with higher insolation. Higher light levels on the windward side during critical growth periods, which also coincide with seasonal storms, may permit kelp to regenerate faster there. However, light is also correlated with factors related to the island's topography making its influence difficult to isolate.
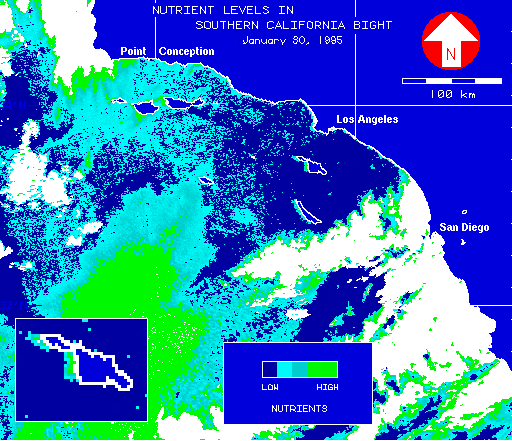
The spatial distribution of kelp, and its ability to recover from disturbance, may be affected by the availability of nutrients such as nitrates (NO-3). Large scale field-measured nitrate levels are not economically feasible for a region as large as the nearshore environment of Catalina Island, and there are no remotely-sensed sources of such data. A strong correlation between temperature and nitrate levels (Zimmerman and Kremer 1984) allowed SST to be used as a surrogate for NO-3 levels. Temperatures in SST images were color-encoded to represent physiologically significant levels. Increasing shades of green represented regions of adequate nutrients; shades of blue-green, borderline levels; and shades of blue, depleted nutrients but acceptable temperatures. Due to the strong correlation between SST and nitrate levels, the inferred patterns of nitrate distribution follow those for ocean temperature. Warmer temperatures on the leeward side result in lower nutrient levels, often to the point of depletion, whereas colder water on the windward side means nutrients may exist at higher levels (figure 22). This suggests that higher nutrient levels on the windward side may also enhance kelp's persistence, even in the face of greater disturbance.
The results of this research substantiate the value of GIS in looking at larger-scale ecological phenomena in the marine environment. Field experiments usually entail limited geographic and temporal scope due to logistics problems, limited time or funding, and the data volumes required. GIS is an appropriate tool for addressing large-scale issues, and an excellent means to identify issues and locations for more intensive field work.
The specific findings in this study indicate the physical complexity and diversity of nearshore marine habitats surrounding Santa Catalina Island may be of great importance in determining the ability of Macrocystis pyrifera to persist. Regionally, distinct differences in storm exposure between the windward and leeward sides, result in large-scale differences in the disturbance regime experienced by kelp. Locally, suitable refugia allowing resistance to disturbance may be found in protected sites with aspects sheltered from direct storm impact, on shallower slopes where wave energy is dissipated more gradually, at deeper depths where wave energy is felt less intensely, or in regions of higher bottom relief which may also offer some protection. On those coasts where disturbance level is high, increased levels of light and nutrients, may contribute to the resilience of populations by enhancing their ability to repair damaged tissue or successfully recruit.
The kelp beds surrounding the island should be viewed as a metapopulation, or population of populations (sensu Levins 1970). The geographic differentiation of persistence has important consequences for metapopulation characteristics, and therefore the designation of appropriate marine reserves to protect both giant kelp and the many marine species that utilize it for habitat or food. Disturbance events may decimate or remove protected kelp beds if they are restricted to a limited range of site conditions, as is presently the case around Catalina. New marine reserves should be designated on the windward side to complement existing ones on the leeward side. Then, in the event of large-scale disturbance such as elevated El Nino temperatures, "remnant patches" (sensu Forman and Godron 1981) of kelp persisting in refugia may provide "seed stock" for the re-establishment of populations around the island.

The author would like to acknowledge the following: the Santa Catalina Island Conservancy and the Offield Family Foundation for their support in the development of several elements in this GIS; Esri for their kind donation of software to the Conservancy which helped make the project possible; SPOT Image Corporation and Dr. Thomas Belsher of IFREMER (France) for the SPOT satellite imagery; Albert Lin, GIS Specialist, Southern California Edison. for the contribution of kelp data layers in ArcInfo format and discussions regarding methods for presenting kelp persistence data; William O'Reilly for his re-creation of the March 12, 1995, peak storm swell image; Rich Cosgrove and Mary Hollinger of NOAA's CoastWatch program for access to AVHRR sea surface temperature data; Dr. M. Neushul, Dr. A. Ebeling and Dr. L. Mertes for their encouragement and advice along the way; Janet Takara for her monumental patience; and many other "anonymous" participants in various INTERNET mail lists and news forums, including Esri-L, GIS-L and comp.infosystems.gis who, unwittingly and unacknowledged, helped through their advice.
Abbott, I. and G. Hollenberg 1976. Marine Algae of California. Stanford University Press. Stanford, California. 827 pp.
Ambrose, R. F., J. M. Engle, J. A. Coyer and B. V. Nelson, 1993. Changes in urchin and kelp densities at Anacapa Island. In Hochberg, F. G. (ed.), Proc. of the 3rd California Islands Symposium: Recent Advances in Research on the California Islands. Santa Barbara Museum of Natural History (Santa Barbara, CA). pp. 199-209.
Bazzaz, F. A. 1983. Characteristics of populations in relation to disturbance in natural and man-modified ecosystems. In Mooney, H. A. and M. Godron (ed), Disturbances and Ecosystems, Components of Response. Springer-Verlag (New York, NY). pp 259-75.
Bushing, W. W., 1994. The influence of topography on the distribution of giant kelp (Macrocystis pyrifera) beds around Santa Catalina Island using a geographic information system. In W. L. Halvorson and G. J. Maender (eds.), The Fourth California Islands Symposium: Update on the Status of Resources. Santa Barbara Museum of Natural History Santa Barbara, CA). pp. 70-82.
Bushing, W. W., 1995. The use of satellite remote sensing and a geographic information system (GIS) to characterize regions of persistent giant kelp (Macrocystis pyrifera) around Santa Catalina Island for designation as marine reserves. Unpublished PhD dissertation. University of California, Santa Barbara.
Connell, J. H. and W. P. Sousa, 1983. On the evidence needed to judge ecological stability or persistence. Amer. Nat. 121:789-824.
Dayton, P. K. and M. J. Tegner 1984. Catastrophic storms, El Nino and patch stability in a southern California kelp community. Science 224:283-285.
Dayton, P. K., M. J. Tegner, P. E. Parnell and P. B. Edwards, 1992. Temporal and spatial patterns of disturbance and recovery in a kelp forest community. Ecological Monographs 62(3):421-445.
Earle, S. 1980. Undersea world of a kelp forest. National Geographic, September, 1980. pp 411-426.
Engle, J. M., 1994. Perspectives on the structure and dynamics of nearshore marine assemblages of the California Channel Islands. In W. L. Halvorson and G. J. Maender (eds.), The Fourth California Islands Symposium: Update on the Status of Resources. Santa Barbara Museum of Natural History (Santa Barbara, CA). pp. 13-26.
Forman, R. T. T. and M. Godron, 1981. Patches and structural components for a landscape ecology. Bio Science 31(10):733-740.
Forman, R. T. T. and M. Godron, 1983. Landscape modification and changing ecological characteristics. In Mooney, H. A. and M. Godron (ed), Disturbances and Ecosystems, Components of Response. Springer-Verlag (New York, NY). pp 12-28.
Foster, M. and D. Schiel, 1985. The ecology of giant kelp forests in California: a community profile. U.S. Fish and Wildlife Service Biological Report 85 (7.2). 152 pp.
Gerard, V. A., 1984. Physiological effects of El Nino on giant kelp in southern California. Mar. Biol. Lett. 5:317-322.
Gore, J. A., J. R. Kelly and J. D. Yount, 1990. Application of ecological theory to determining recovery potential of disturbed lotic ecosystems: research needs and priorities. Environmental Management 14(5):755-762.
Hagen, N. T., 1983. Destructive grazing of kelp beds by sea urchins in Vestfjorden, Northern Norway. Sarsia 68:177-190.
Harger, B. W. 1983. A historical overview of kelp in southern California. In Bascom, W. (ed) The Effects of Waste Disposal on Kelp Communities. California Sea Grant Symposium. 328 pp.
Harrold, C. and D. Reed, 1985. Food availability, sea urchin grazing, and kelp forest community structure. Ecology 66:1160-1169.
Hodder, D. and M. Mel, 1978. Kelp survey of the Southern California Bight. Esca-Tech Corp. and Science Applications, Inc. Technical Report Volume III - Report 1.4 to the Bureau of Land Management (Year II SCOCS Program), Contract No. AA550-CT6-40, La Jolla. 104 pp.
Holland, P. G. and D. G. Steyn, 1975. Vegetational responses to latitudinal variations in slope angle and aspect. Journal of Biogeography 2:179-183.
Jackson, G. A., 1977. Nutrients and production of giant kelp, Macrocystis pyrifera, off southern California. Limnol. Oceanogr. 22:979-995.
Kennelly, S. J. and A. J. Underwood, 1993. Geographic consistencies of effects of experimental physical disturbance on understory species in sublittoral kelp forests in central New South Wales. J. Exp. Mar. Biol. Ecol. 168(1):35-58.
Krcho, J., 1991. Georelief as a subsystem of landscape and the influence of morphometric parameters of georelief on spatial differentiation of landscape-ecological parameters. Ekologia (CSFR) 10(2):115-57.
Lawrence, J. M., 1975. On the relationships between marine plants and sea urchins. Ann. Rev. Oceanogr. Mar. Biol. 13:213-286.
Leighton, D. L., L. G. Jones and W. J. North, 1966. Ecological relationships between giant kelp and sea urchins in southern California. Proc. Intl. Seaweed Symp. 5:141-153.
Levins, R. 1970. Extinction. Lectures in Mathematics in the Life Sciences 2:75-107.
Mann, K. H. and P. A. Breen, 1972. The relationship between lobster abundance, sea urchins and kelp beds. J. Fish. Res. Board Can. 29:603-605.
McAlary, F. A. and W. N. McFarland, 1994. Catalina Island kelp forests: 1992-1993. In W. L. Halvorson and G. J. Maender (eds.), The Fourth California Islands Symposium: Update on the Status of Resources. Santa Barbara Museum of Natural History (Santa Barbara, CA). pp. 35-44.
Neushul Mariculture, Inc. 1981. Historical Review of Kelp Beds in the Southern California Bight. Southern California Edison Company Research Report Series Number 81-RD-98. 74 pp.
North, W. J. 1971. The biology of giant kelp beds (Macrocystis) in California. Beihefte Zur Nova Hedwigia, Vol. 32. 600 pp.
North, W. J. 1991. The Kelp Beds of San Diego and Orange Counties. Published by Wheeler J. North, March 15, 1991. 270 pp.
North, W. J. and J. S. Pearse, 1970. Sea urchin population explosion in southern California coastal waters. Science 167:209.
Norton, J., D. McLain, R. Brainard and D. Husby, 1985. The 1982-83 El Nino event off Baja and Alta California and its ocean climate context. In Wooster, W. S. and D. L. Fluharty (eds.), 1985. El Nino North: Nino Effects in the Eastern Subarctic Pacific Ocean. Washington Sea Grant Program, University of Washington (Seattle, WA). pp. 44-72.
O'Reilly, W. C. and R. T. Guza, 1993. A comparison of two spectral wave models in the southern California Bight. Coastal Engineering 19:263-282.
Pearsons, T. N. and H. W. Li, 1992. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Amer. Fish. Soc. 121:427-436.
Pond, S. and G. L. Pickard, 1983. Introductory Dynamical Oceanography. Pergamon Press (New York, NY). 329 pp.
Quast, J. C. 1971. Some physical aspects of the inshore environment, particularly as it affects kelp bed fishes. In W. J. North (ed.), The Biology of Giant Kelp Beds (Macrocystis) in California. Beihefte Zur Nova Hedwigia 32:228-240.
Rabinowitz, D., J. K. Rapp, S. Cairns and M. Mayer, 1989. The persistence of rare prairie grasses in Missouri: environmental variation buffered by reproductive output of sparse species. The Amer. Natur. 134(4):525-544.
Reed, D. C. and M. S. Foster 1984. The effects of canopy shading on algal recruitment and growth in a giant kelp forest. Ecology 65(3):937-948.
Reice, S. R., R. C. Wissmar and R. J. Naiman, 1990. Disturbance regimes, resilience, and recovery in animal communities and habitats in lotic ecosystems. Environmental Management 14(5):647-659.
Richards, D. V., D. Kushner and W. Avery, 1993. Kelp forest monitoring- Channel Islands National Park (1990 Annual Report). Technical Report NPS/ERUC/NRTR-93/05. National Park Service. University of California (Davis, CA). 195 pp.
Ripple, W. J., G. A. Bradshaw and T. A. Spies, 1991. Measuring forest landscape patterns in the Cascade Range of Oregon, USA. Biological Conservation 57(1991):73-88.
Santelices, B. and F. P. Ojeda, 1984. Effects of canopy removal on the understory algal community structure of coastal forests of Macrocystis pyrifera from southern South America. Mar. Ecol.-Progr. Ser. 14(2):165-173.
Seymour, R., M. Tegner, P. Dayton and P. Parnell 1989. Storm wave induced mortality of giant kelp, Macrocystis pyrifera, in southern California. Estuarine, Coastal and Shelf Science 28:277-292.
Sousa, W. P., 1984. The role of disturbance in natural communities. Ann. Rev. Ecol. Syst. 15:353-391.
State Water Resources Control Board 1979. California Marine Waters: Areas of Special Biological Significance, Reconnaissance Survey Report, Santa Catalina Island- Subarea I, Los Angeles County. Water Quality Monitoring Report No. 79-6. April, 1979. State Water Resources Control Board, Division of Planning and Research, Surveillance and Monitoring Section. 192 pp.
State Water Resources Control Board, 1981. California Marine Waters: Areas of Special Biological Significance Reconnaissance Survey Report, Santa Catalina Island- Subarea II, North End of Little Harbor to Ben Weston Point and Subarea IV Binnacle Rock to Jewfish Point, Los Angeles County. State Water Resources Control Board, Division of Planning and Research, Surveillance and Monitoring Section. Water Quality Monitoring Report No. 81-10. October, 1981. 190 pp.
Steinman, A. D., P. J. Mulholland, A. V. Palumbo, T. F. Flum and D. L. DeAngelis, 1991. Resilience of lotic ecosystems to a light-elimination disturbance. Ecology 72(4):1299-1313.
Stevens, J., 1994. Twenty year database of fish off Los Angeles. Emeritus professor, Occidental College, Los Angeles. Seminar talk at the University of California, Santa Barbara, May 2, 1994.
Tegner, M. J., 1980. Multispecies considerations of resource management in southern California kelp beds. In J. D. Pringle, G. J. Sharp and J. F. Caddy (eds), Proc. of the Workshop on the Relationships Between Sea Urchin Grazing and Commercial Plant/Animal Harvesting. Canadian Technical Report, Fisheries and Aquatic Sciences 954. pp. 125-143.
Weaver, M. and M. Kellman, 1981. The effects of forest fragmentation on woodlot tree biotas in Southern Ontario. Jour. of Biogeography 8(1981):199-210.