Analyzing Wildlife Movement Corridors in Montana Using GIS
Richard Walker and Lance Craighead
Abstract
We delineated landscape routes offering the best chance of success for wildlife moving among the three large core protected areas in the Northern Rockies -- the Salmon-Selway, Northern Continental Divide, and Greater Yellowstone Ecosystems. Using ARC/GRID and Montana Gap Analysis data, we derived habitat suitability models for three umbrella species, which we combined with road density information to create kilometer-scale cost surfaces of movement. For each of the three species -- grizzly bear, elk, and cougar -- we performed a least.cost.path analysis to locate broad potential corridor routes. From this first approximation we identified probable movement routes and as well as critical barriers, bottlenecks, and filters where corridor routes intersected with high.risk habitat. This analysis is being used to identify priority areas for wildlife management to improve the connectivity between the core protected ecosystems in the Northern Rockies.
Introduction
Habitat reduction and fragmentation at a variety of spatial scales has been widely acknowledged as a primary cause of the decline of many species worldwide (Ehrlich 1986, Lovejoy et al. 1986, Harris 1984). Habitat fragmentation generally leads to smaller and more isolated animal populations. Smaller populations are then more vulnerable to local extinction, due to stochastic events (Shaffer 1978, Gilpin and Soule 1986), and they are more susceptible to the negative effects of inbreeding depression. To reduce the isolation of habitat fragments, many conservation biologists (e.g. Noss 1983, 1987, Noss and Harris 1986, Craighead et al. 1997, Craighead and Vyse 1995, Paetkau et. al. 1997) have recommended maintaining landscape "connectivity" -- preserving habitat for movement of species between remaining fragments.
While a significant body of research has demonstrated the deleterious effects of habitat fragmentation, work examining critical thresholds of habitat connectivity has lagged behind. For several species, Population Viability Analysis (PVA) has been employed to estimate minimum viable populations as well as the amount of habitat necessary to support a minimum viable population (MVP) (Shaffer 1978, 1981, 1987, Gilpin and Soule 1986). Yet few similar analyses have been done explicitly to demonstrate minimum thresholds of connectivity for adequate protection of species. The scientific debate over the efficacy or even necessity of species corridors is a reflection of this problem (Simberloff and Abele 1976, Simberloff et al. 1992). However, since habitat fragmentation has been shown to contribute to population declines, by inference some degree of "lack of fragmentation", or habitat connectivity, should help ensure long-term species survival (Noss 1987, 1992, Beier 1993, Noss et al. 1996).
At regional scales, connecting large core areas of wildlife habitat requires corridors -- land managed for its function as routes for wildlife movement and dispersal (Saunders and Hobbs 1991). The notion of connective habitat corridors implies a system of corridors and the core areas of habitat which they serve to link. Conceptual models of core areas, movement corridors and buffer zones have been proposed by several workers (Soule, Noss 1992, Noss and Harris 1996) as frameworks for long-term regional scale conservation of wildlife.
The Ninth U.S. Circuit Court Of Appeals (1990) provided a reasonable definition
of a corridor:
"...avenues along which wide-ranging animals can travel,
plants can propagate, genetic interchange can occur,
populations can move in response to environmental changes
and natural disasters, and threatened
species can be replenished from other areas."
Presently, corridor or connectivity analysis has been mandated in the Upper Columbia River Basin Integrated Scientific Assessment (Quigley et al. 1996), in the U.S. Forest Service conservation strategy for Western forest carnivores (Ruggiero et al. 1994), and in the Interagency Grizzly Bear Recovery Plan (USFWS 1993). Corridor analysis is a logical tool for compliance with the letter and spirit of the National Forest Management Act of 1976, the Endangered Species Act of 1973, and the National Environmental Policy Act of 1972. Wildlife movement corridors have been mentioned or broadly outlined in Environmental Assessments such as Montana Department of Fish, Wildlife and Parks conservation easements for the Sieben Ranch (MDFWP 1996) and Lewis and Clark National Forest Oil and Gas Leasing documents (USDA 1996). Despite this recent acknowledgment of the importance of linkage zones, the only guidelines so far provided by government agencies is a list of 8 types of information layers to be used in a GIS for linkage zone assessment, described in the grizzly bear recovery plan. A 5-year process of regional-scale linkage assessment for grizzly bear populations in the lower 48 states is currently underway (USFWS 1993).
In order to contribute to defining a scientific framework for linkage zone assessment (or corridor analysis) we report an independent effort to delineate wildlife corridors according to the conservation biology model (cores, corridors, buffers) at regional scale in a specific geographic area. Using Geographic Information System (GIS) software, the best available spatial data on habitats, and considering the habitat preferences of 3 select umbrella species, we have modeled potential regional-scale wildlife corridors between core protected areas in the Northern Rockies of the United States. Our approach offers a biologically defensible assessment of probable corridor routes and suggests one means (least-cost) of estimating the relative "connectivity" of alternate routes. A flowchart of the process we follow looks like this:
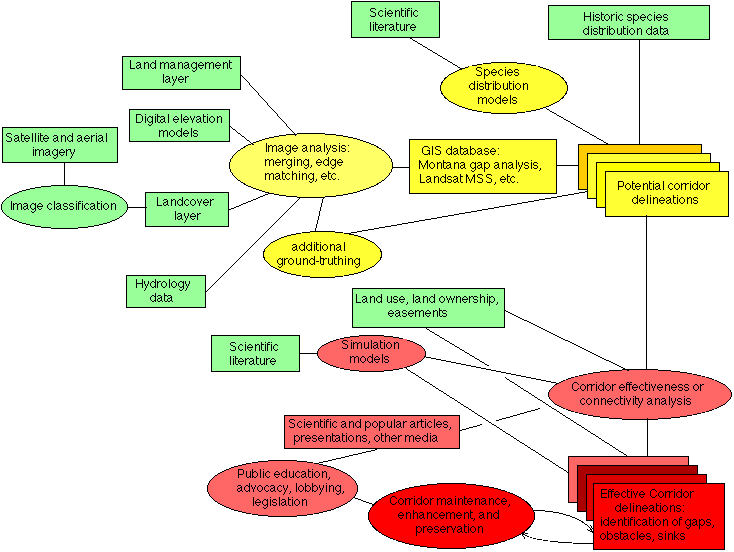
where rectangles represent data sources and products, and ovals represent processes or techniques. Green items were available at the beginning of the project, Yellow items are the subject of this paper, and Red items are further refinements of the analysis and implementation steps necessary to protect corridor habitat.
Background
The Northern Rockies of the United States, comprised primarily of western Montana, central and northern Idaho and northwestern Wyoming, harbor some of the last vestiges of North Americaąs great biological heritage. Here are the last remaining populations of grizzly bears and free-ranging bison. In several areas, the full complement of large native predators present at the arrival of Columbus in the New World still persists.
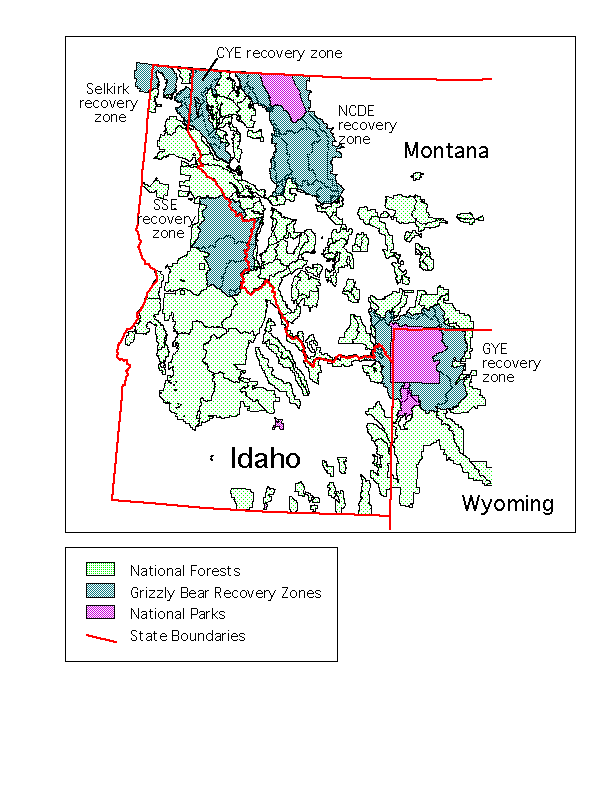
With increasing human development, however, wildlife habitat in the region is becoming ever more fragmented. New roads, housing developments, and natural resource extraction activities have caused major changes in the natural landscapes over the past few decades, and in the process have removed or isolated areas of habitat formerly available to wildlife. Projections are for this trend of habitat fragmentation to continue and accelerate, as the Northern Rockies is one of the fastest growing regions in the country (Quigley et al. 1996).
One result of the regional scale fragmentation in the Northern Rockies is the current situation of the grizzly bear, which is now isolated in a handful of remnant disjunct populations. The bear populations are centered in large, relatively undeveloped and undisturbed areas including the Greater Yellowstone Ecosystem, the Northern Continental Divide Ecosystem and, to a much lesser degree, in the mountains of northern Idaho and northwest Montana (USFWS 1993). A process has been initiated to restore the grizzly to the Salmon Selway area in central Idaho (USFWS 1996).
The grizzly bear is a sensitive regional indicator of fragmentation and the effects of human development. It is considered an "umbrella species" -- a species whose proper long-term management would likely help to ensure the persistence of many other species which also occur in the ecosystem. Because they require large areas of relatively undisturbed habitat, solving for the habitat requirements of grizzlies can assist in defining large core protected areas, and smaller protected areas to serve as nodes in a networked regional landscape habitat system.
Cougars, which are widespread in the Northern Rockies, have fairly well-defined habitat requirements, including forest cover, forest edge and prey. Conserving habitat for the regional movements of cougars, much as for the grizzly bear, would also likely provide protection for other mammals moving through a landscape. In addition, we also examined and modeled the habitat needs of elk for dispersal. Protecting adequate corridors for these three species -- a large carnivore, a large omnivore and a large ungulate -- is also likely to provide a protective umbrella for a number of other smaller inhabitants of the region. The following discussion is in the general context of these three umbrella species.
Dispersal behavior
Dispersal patterns of subadult animals as they reach sexual maturity are an important factor determining gene flow, population genetic structure, and ultimately genetic diversity in populations. Animals dispersing from "source" populations can be recruited into declining or "sink" populations to maintain demographic stability. In theory, any animal in a population can disperse away from its original home range. Research, however, has shown a tendency for most dispersing animals to fit a more limited profile. In mammals, the general pattern of dispersal seems to be that males are more mobile, while females remain nearer to their natal areas (Greenwood 1980). Understanding the behavioral characteristics of those animals most likely to disperse can assist in determining criteria for assessing the best routes for corridor habitat protection, since these animals are also the most likely to use and benefit from corridors.
In the case of large predators such as grizzly bears and cougars, the most typical dispersing animal is a juvenile male. Maternal females will not tolerate the presence of their juvenile male offspring in their home ranges, and thus force them away at a certain age. In many cases, the surrounding habitat is occupied by older, more dominant males. Thus a banished juvenile male often has no choice but to disperse to a different area as yet unknown to him, and search for adequate prey and cover, and a territory he can occupy. It is generally hypothesized that dispersing animals will occupy the first unoccupied habitat they encounter (Waser 1985). However, there are instances where dispersers are tolerated within adult home ranges (McCord 1974), and other instances where dispersers travel long distances (sometimes returning later) even though suitable habitat appeared to be found nearby (Craighead 1994).
Dispersing animals are typically young and relatively inexperienced. In addition, they are moving through unfamiliar landscapes, which can lead to some random exploratory behavior and less predictable patterns of movement. As they wander through terra incognita, they may tend to avoid certain phenomena and be attracted to others, and can be influenced by factors such as scent and wind direction that are not obvious to humans. To some degree, their movements may be indistinguishable from random.
For our purposes, delineating corridors at regional scale entailed determining which routes, based upon the observed needs of wildlife, offer an animal the best chance of survival if it were to disperse between core protected areas. In this effort we have attempted to balance two general factors affecting wildlife movements: the most suitable connected habitat (in the absence of humans) and the degree to which human-related impediments are present in the landscape.
We do not make the claim that these routes are most the likely to be used by dispersing individuals. Due to highly variable individual behavior this would be very a tenuous assumption. Rather, our analyses indicate that other routes, while perhaps equally likely to be taken by an individual, would reduce his/her probability of survival in the process of dispersing from one core protected area to another, generally because of increased transit time and risk of encountering significant impediments.
Thus the route delineations from our analyses are more "prescriptive" rather than strictly "descriptive". The objective of protecting such corridors would be to leave the best routes open to wildlife, but without implying any mechanism of forcing animals to choose those routes (other than the fact that increasing development and change outside of potential corridors will continually make those areas more and more hostile to most wildlife species).
Methods
Model Assumptions
Wildlife respond to landscapes (and habitat) at several scales. Foraging and hunting activities, for example, can be geared to microsite occurrences of vegetation or prey (for example see Apps 1996). We assumed that dispersing animals are less sensitive to local environments, and respond to a larger landscape in their movements. For modeling best regional scale corridor routes we made the following assumptions.
Assumption 1) Good corridors are comprised primarily of preferred habitat types
Many sorts of habitats can allow for wildlife movement, even those which are not generally preferred. In one sense, large open areas can facilitate movement, in that the terrain offers few impediments to transit. On the other hand, many species avoid open areas lacking adequate vegetation cover for concealment. While it is possible that "good transiting habitat" is not necessarily the same as habitat preferred for most other functions (e.g. hunting, grazing, bedding, etc.), in our work we assumed that corridors should be comprised primarily of generally preferred habitat types. Such habitat offers a greater probability of at least short-term survival for an animal, given that it is more likely to be able to find adequate food and shelter therein. However, species such as those we model could pass through unsuitable habitat for one to several days without significant harm.
Assumption 2) Humans pose problems for successful transit
In human-dominated landscapes, particularly during hunting season, wildlife species learn to either avoid humans and developments, or they learn to associate humans with sources of food. The problems humans pose for transiting wildlife can be broken down into two types.
First, their dwellings can attract dispersing individuals, through easy access to food (e.g. pet food on the porch, poultry feed, garbage). Inexperienced animals can easily become habituated to humans, and cause increasing problems. Habituated individuals are more likely to be killed or transported to some remote locale, thus terminating their dispersal through that area.
Secondly, in many areas human activities can repel these "wary" transiting individuals, by loud machine noise or other high profile activities. They can often displace wildlife from important habitat and force them to use other, less-valuable habitat. It can also effectively restrict their moving to otherwise available habitat in nearby areas, reducing the chances of survival for the individual and ultimately the population.
People also have a general tendency, when encountering large predators, to kill them directly. Because of the problems many species encounter with humans, optimal corridor routes should avoid human developments and activities as much as possible.
Assumption 3) Current human developments are permanent
Closing or destroying some key human developments could greatly change the value of particular corridor route. However, for this initial analysis we assumed that all developments, including roads, are permanent. Subsequent analyses may include alternate scenarios -- i.e. what effect overall corridor ratings would result from closing roads or altering land management practices.
Assumption 4) Least-cost path
We assumed that the least-cost path, based upon the previous assumptions, offers an animal the greatest probability of survival in traversing the entire distance. This was facilitated by ArcInfo GRID functions costdistance and corridor, but it also has a significant degree of biological relevance. Although a dispersing animal may not choose the least-cost (optimum) path, if it did it would encounter fewer hazards or obstacles, would spend less time in traveling, and would travel through habitat with higher probability of containing food and concealment cover, thus increasing its probability of survival. The least-cost path balances habitat suitability, minimum euclidean distance and degree of "connectivity" between the two endpoints.
GIS Model inputs
We developed three input GIS coverages to model best potential corridor routes: 1) habitat quality (per species per vegetation type); 2) amount (length) of forest and shrub/grassland interface; and 3) road density. The first two are measures of the quality of an area in terms of its utility to a species, while the third is a good indicator of the amount of human use and disturbance in the area. These measures are similar to the parameters developed for the computer-based Cumulative Effects Model for the Grizzly Bear, by researchers with the Interagency Grizzly Bear Committee (Systems for Environmental Management, 1994), and others.
Habitat quality
The Montana Gap Analysis project recently produced detailed GIS coverages of dominant vegetation cover for most of western Montana and central and northern Idaho. Several gap analysis coverages relate to vegetation cover. For our habitat quality model we used a raster (cell) version of the gap analysis vegetation cover types. This file maps more than 50 different cover types, including some which were not native vegetation (e.g. barren, agriculture). Based upon an extensive review of the literature, we then rated each vegetation type according to its quality for grizzly bear, cougar and elk. Rating values ranged from 0 (unsuitable) to 3 (highly preferred) for each species.
Habitat quality, however, is not strictly a function of the presence of preferred types. The spatial pattern of type mixtures can enhance or detract from the overall habitat quality. Grizzly bears, cougar and elk are known to prefer a mixture of cover (for security) and open areas (for food or prey) over singular forest or grassland types. To indicate this "metatype" quality of habitat mixes, we derived the length of forest/grassland and forest/shrubland boundaries for the Gap analysis region.
Roads data
As a measure of human development and use, we at first used the predicted road density coverage of the Interior Columbia Basin Ecosystem Management Project (ICBEMP), which covers most of the region. After working with the coverage, however, many errors and inaccuracies became obvious. The data were clearly of insufficient quality to use in the analyses. Instead, we extracted roads information from two digital sources -- U.S. Census Bureau TIGER files and U.S. Forest Service Cartographic Feature files. While some inconsistencies in the source files were evident, overall the quality of the data were high. (The 1:100K scale USGS Digital Line Graph data was of similar quality to the TIGER files, but were more problematical to use).
Not all roads have equal impact on wildlife and the landscape. We thus weighted the roads according to their estimated use. In the TIGER files, all major highways were given a weight of 3, other major roads a weight of 2, and all other roads and railroads a weight of unity. The weights are roughly proportional to amount of disturbance or degree of difficulty an animal might have in attempting to cross the road.
Integrating landscape variables at regional scale
A central question in modeling wildlife habitat and corridors is determining the best scale, or set of scales, at which to perform the analyses. Many ecological studies demonstrate that animals learn and "map" their home range areas and know where food, cover and other requirements are located on a "micro-scale" that can often be measured at a scale as fine as square meters. However, animals such as elk, cougar and grizzly bear regularly move over long distances through habitat that is not of any particular value to them in order to reach areas they prefer. In addition, dispersing individuals may wander for days through poor habitat before encountering better habitat where they can find food.
For regional scale corridor routes, estimating habitat variables at the scale of 30 meters (the resolution of Landsat Thematic Mapper and Montana Gap Analysis) cannot be supported by the level of detail of the data. A model using data of such fine scale would be highly sensitive to local small changes in suitability rating, and may be unrealistic in its results. Thus, to examine possible regional-scale movement routes, given the limitations of GIS data and habitat characterization, we integrated habitat quality and roads data over larger areas. The choice of area unit size was critical -- too coarse a scale could obscure important variations in suitability of the landscape for a wildlife corridor.
For this analysis we integrated the landscape (and habitat) variables at a resolution of one square kilometer. This scale offered a reasonable balance between the fine scaled information base (30m in the case of habitat data, continuous in the case of roads data) and a broader scale unit (e.g. a small watershed). Ignoring the filtering effect of the 2.5 hectare minimum mapping unit of the Montana Gap Analysis database, integrating the habitat value coverage involved averaging approximately 1110 30m cells per square kilometer. The reasoning behind the selection of square kilometer integrations is further elaborated upon in the following discussion.
UTM-based square kilometer integrations
The information from Gap Analysis were derived from 30 meter cells, which had been generalized into minimum patch sizes (mapping units) of 5 acres (2 hectares). To provide a spatial integration of the 30m cells, we generated a square kilometer grid overlay based upon the Universal Transverse Mercator (UTM) coordinate system. . The square kilometer cells conform to 1000m increments in the regional UTM zone (i.e. each cell is bounded by whole kilometers in projected coordinates).
We selected the UTM square kilometer cell integration for several reasons. First, it provided a filter of the landscape in line with a characteristic scale of larger wildlife sensory perception, on order of a 500m radius (this is, of necessity, a generalization -- scents and sounds can easily carry for several kilometers). Second, it offered convenience in the field, as UTM grids overlay all recently revised USGS 1:24000 series maps. Third, global positioning system (GPS) units conventionally display ground coordinates in the UTM system. Had we employed an arbitrary set of integrating cells of the same kilometer resolution, the our ability to assess the accuracy of our work would have been compromised, as it would have been much more difficult to assess the limits (and thus accuracy) of such cells in the field.
Based upon the literature and expert opinion, we derived mean habitat quality values for each UTM-based square kilometer cell for grizzly bear, cougar and elk. The mean values varied from 0 to 3. For each cell we also derived the total length of forest/shrubland edge and forest/grassland interface. These values ranged from 0 to about 11 kilometers per square kilometer. Though they are not strictly independent, the habitat value and edge length coverages provided us with two measures of the suitability of each square kilometer for these species.
Road density was obtained in a similar manner to the forest edge coverage. Roads were assigned weightings according to their approximated traffic load, then the weighted length totals were derived for each UTM cell. The range of the cells was very wide -- from 0km/km2 in roadless and wilderness areas to nearly 20km/km2 in highly urbanized areas.
Synthesizing the input parameters
For each of the three species, the three landscape variables (habitat quality, forest edge and road density) required synthesis into a single coverage layer. This matrix represented a model landscape "surface" offering varying degrees of resistance to movement per species per cell. The higher the resistance or "cost" associated with crossing the cell, given the context of its surrounding cells, the less important or suitable the cell was apt to be as part of a corridor.
Combining the three variables to derive overall habitat suitability required some notions of the relative importance of the factors and the way in which they interrelate. As a springboard for our modeling, we used both the relative ranges of the variables and a functional form similar to that of the Cumulative Effects Model developed for the grizzly bear (SEM, 1994). In that still-evolving model, the "vegetation coefficient" (i.e. our habitat value) and "disturbance coefficient" (analogous to our road density parameter) varied between 0 and 1, while the "edge coefficient" (i.e. edge length) and "supplemental coefficient" were normalized to values between 0 and 2. The general form of the (atemporal) CEM ultimately computes habitat effectiveness parameter via the following:
Habitat Value = (vegetation coefficient) * (edge coefficient) *
(supplemental coefficient) * (disturbance coefficient)
Our model resembled the form of the CEM, but produced a value inversely proportional to the suitability of the cell for a corridor. Specifically, it had the following form:
Cell Resistance = -1.0 * ((habitat coefficient) * (edge coefficient)) +
(road density coefficient)
For the edge and the road density coefficients, we took log transforms of the original values (units of km/km2), then normalized them to the desired maximum values. This reduced the sensitivity of the model to extreme values of road density and edges, and increased the sensitivity to changes in the lower ranged values.
The habitat value and edge coefficients clearly interact and are not entirely independent measures. Thus we multiplied these two terms together. The road density was independent of the other factors, thus we added it to the other terms.
For elk and grizzly bear we used the above form of the model. For cougar modeling, since little is reported in the literature on road/human sensitivity, we omitted the road density coefficients from the calculations.
Results
Using ArcInfo GRID functions costdistance and corridor, with our species-specific movement "cost surfaces" as inputs, we created symmetrical cumulative cost surfaces with pairs of core protected areas of the Northern Rockies as origins/destinations. For the core areas we took units managed specifically for wilderness on public lands which were contiguous within an ecosystem. The Salmon-Selway Ecosystem core area was comprised of all contiguous wilderness areas in central Idaho, including the Gospel Hump, Selway-Bitterroot and Frank Church/River of No Return wilderness areas. For the Greater Yellowstone Ecosystem, Yellowstone National Park and all National Forest Wilderness Areas contiguous with the park were used. The Northern Continental Divide Ecosystem was comprised of Glacier National Park and the Bob Marshall wilderness complex.
Least-cost travel grids of were generated for each combination of the three species and three core protected areas. These consisted of cumulative distances calculated for every cell in the region from the nearest point of a given defined core protected area, weighted by the habitat suitability cost grid.
The best corridors indicated between a pair of protected areas varied according to the species, as may be expected given differing habitat preferences and sensitivity to human activities. A brief synopsis of each modeled corridor is given below.
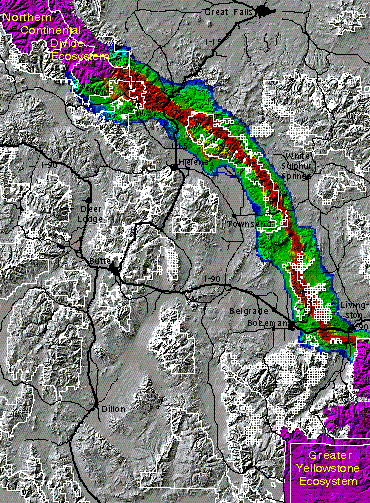
The Greater Yellowstone -- Northern Continental Divide connection
The distance between the Greater Yellowstone Ecosystem (GYE) and the Northern Continental Divide Ecosystem (NCDE) is approximately 200 kilometers. The area in between consists of a complex of forested mountains and open grassland/sagebrush valleys, with varying connectivity among them. The three species analyzed all tend to maintain close proximity to tree cover. They are likely to be unable to use the expanses of wide open grasslands and shrublands which occur in areas such as the Missouri River Valley near Townsend, MT. For all species, any route from the GYE to the NCDE involves crossing two interstate highways -- I-90 and I-15.
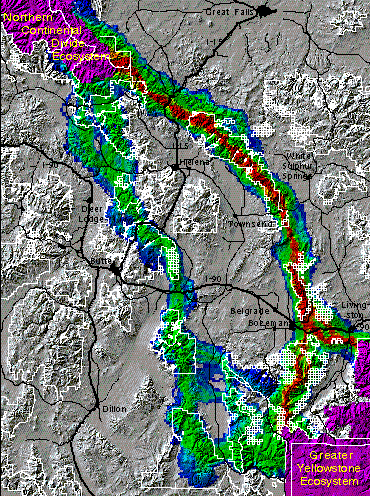
The figure below shows the results of the grizzly bear model for this regional linkage. The corridors in this figure are color-coded, with warmer colors indicating better areas of habitat connectivity. Bottlenecks are indicated in parts of the corridors which are more constricted. The first and second best major corridor routes for grizzly bears are depicted, with one route being superior to others. According to our work, the potential corridor offering the best chance of successful transit consists of the Gallatin, Bridger, and Big Belt mountain ranges. A secondary route for bears, far inferior to the primary in this analysis, is comprised primarily of the Taylor-Hilgard, Gravelly, Tobacco Root, Whitetail/ONeil, and Boulder mountain ranges.
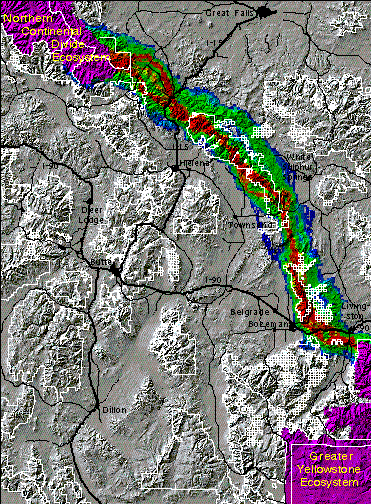
For elk the route which appears to offer the best corridor (shown below) leaves the GYE at the north end of the Absaroka Range and continues to the NCDE via the Bridger and Big Belt ranges. The main corridor routes for elk converge with the primary grizzly route in the Bridger range. The former, however, has several interweaving braids of highly suitable corridor routes, including three which roughly parallel each other between Interstate 90 and the NCDE. Our model indicates more flexibility in corridor planning for elk than for grizzly bears in these areas.
The cougar preferred corridor route (below) also in large part follows the spine of the Big Belt and Bridger ranges. As with the elk, in some areas more than one main route is indicated, although the primary route is in all cases much wider than the others. This results suggests that a single wide corridor may suffice for transiting cougars in linking the GYE with the NCDE.
The Greater Yellowstone -- Salmon-Selway connection
Although we modeled corridors for all three species in this area, we present here only the results for grizzly bears to illustrate one set of potential corridors. Grizzly bears originating in either the GYE or Salmon-Selway Ecosystem (SSE), according to our model, might best transit this roughly 380 kilometer distance along a route comprised mainly of the Centennial Mountains dividing Montana from Idaho. From the east this corridor begins from the south end of the Madison Range, follows nearly 200 kilometers along the continental divide (following the Centennial mountains), and then crosses over the Lemhi Valley to the Lemhi Range of Idaho. From the Lemhi Range two routes fork and head west into the Frank Church - River of No Return Wilderness area.
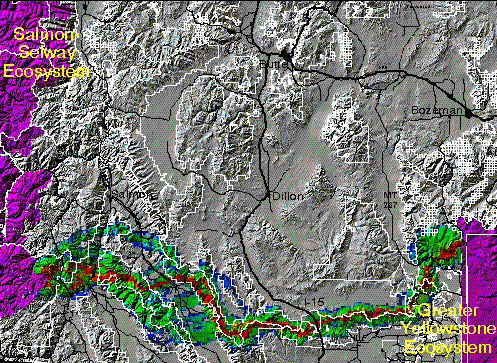
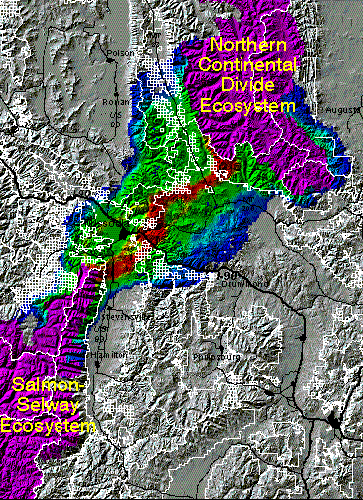
The Salmon-Selway -- Northern Continental Divide connection
The distance between the Salmon-Selway (SSE) and Northern Continental Divide Ecosystems (NCDE) is much shorter in air miles than between the other core areas. The least cost path corridor routes modeled from the SSE to the NCDE were similar for all three species. A representative example is the grizzly bear corridor presented below. Within the constraints of the methods, the best routes crossed from the Bitterroot Mountains to the north end of the Sapphire mountains, then arc to the north, passing west of Rattlesnake wilderness area. The corridor then takes a fairly direct route to the southwest lobe of the Bob Marshall wilderness complex. This route passes close to the densely inhabitat area around Missoula, MT.
Discussion
Although there has been debate over the value of corridors, we feel that the arguments are largely semantic in nature. There can be little doubt that individual animals disperse over long distances from their natal areas and that these movements constitute gene flow and demographic interchange between populations. As habitat becomes fragmented, dispersal routes are cut off or deflected. If a route remains over which an animal can conceivable travel, it may as well be termed a "corridor". Those who are skeptical of the corridor concept are concerned that there is often little empirical evidence that a given corridor may actually function as such for the species in question. On the other hand there are very many instances where animals have been shown to use what little habitat remains in an area for movement. Opponents worry that the cost of providing a corridor may outweigh the benefits. This may be the case in highly developed landscapes where habitat must be restored in expensive real estate in order to provide a corridor, but it is of much less concern in landscapes like the Northern Rockies where the habitat and the connections still exist but are rapidly being altered.
On the basis of radiotracking data and genetic data alone in the Northern Rockies, we are confident that animals disperse on a regional basis. Some species such as wolves or cougars can cover the distance between protected reserves in a single season; others such as grizzly bears, fisher, lynx, or pine marten may take several generations to move from one large reserve to another. We used a first approximation model to define the areas most useful to maintain this connectivity. The results of our regional scale wildlife corridor analyses indicate several routes which may provide the best opportunities for successful animal transits. At this first glance, they appear to be reasonable, as they transit areas known to contain good habitat and which in general lack people.
Results have not yet been validated in the field, although various segments of the routes we arrived at are known to be used as travel routes by various species. In particular, many of the high-risk bottlenecks where animals must cross highways are convergent with our mapping analysis and records of road-kills or radiolocation data.
The results are strongly dependent on the accuracy of the model inputs, as well as on the formula used to combine them into a single dataset (cell resistance to transit). There may be a variety of other formulas which are equally valid. Other than the grizzly bear cumulative effects model, there is little in the literature to help guide the variable weighting and the specific form of the model.
Conclusions
The concept of corridors has become well entrenched in conservation thinking; however, the methodology and implementation of these concepts is only now beginning to take shape. As we move slowly towards a systematic consensus of definitions and techniques, critical habitat for linkage of populations may be altered without an understanding (or in spite of) its importance for wildlife. As the U.S.Fish and Wildlife Service Grizzly Bear Recovery Plan states:
"Future land management activities within these areas may be critical to
maintaining their utility as linkage zones. It is essential that existing options for
carnivore movement between existing ecosystems be maintained while the
evaluation of linkage zones is underway. Management strategies that limit
human-induced mortality and address access management will facilitate the
maintenance of the potential of these zones during the 5-year evaluation period.
On public lands, management prescriptions similar to big game summer range
prescriptions that address access management would likely conserve any existing
potential of these areas for linkage until completion of the 5-year evaluation process."
As of this writing, the 5-year process of linkage assessment for grizzly bears by the U.S. Fish and Wildlife Service will take at least one more year. This analysis is based upon the grizzly bear cumulative effects model (ICE6 1994), a complex habitat suitability model that is nearing completion of validation by each recovery zone. It should adequately address linkage concerns for grizzly bears when it is done: currently it is being applied to a pilot study in the Swan Valley of Montana. However, in the interim there is no mechanism for evaluating potential corridor areas or ensuring that "existing options for carnivore movement between existing ecosystems be maintained while the evaluation of linkage zones is underway". In addition there is no concerted agency effort to address habitat connections for populations of other threatened or sensitive species such as wolverine, fisher, or lynx.
We offer this approach to identifying important habitat connections for wildlife movement in the Northern Rockies as a first-generation, simple, broad-scale picture of existing habitat that has a high probability of connecting the large core reserves for dispersing wildlife species. We feel that the three "umbrella species" we have chosen should serve to delineate core areas and buffer zones for movement corridors that will include habitat needs of the majority of wildlife species. We hope that land management agencies will incorporate the results of our analysis in planning management activities within the areas we have identified as probable corridors, and postpone any significant alteration of existing habitat until more refined and more comprehensive models are developed.
Future work
The least-cost path analysis is one of several techniques for indicating best routes for wildlife through the Northern Rockies. It offers advantages in terms of ease of computation and interpretation. A disadvantage is that it may not be the path of least resistance -- i.e. the path through which the average cell (resistance) value may be minimized, in which a longer route could be superior. Delineating a 'least average cost' path, however, would be much more computationally demanding, due to the number of possible cell-path combinations in a large region.
Another method we are pursuing is a Monte Carlo simulation in which we are modeling the three species movements in the same 'resistance to transit' cell grids, given varying degrees of randomness. The results of this type of analysis may provide a more realistic view of corridors, bottlenecks and other impediments than the strictly deterministic least-cost path analysis described in this paper.
Literature Cited
Apps, C., 1996. Bobcat (Lynx rufus) habitat selection and suitability assessment in southeast British Columbia. M.S. Thesis. University of Calgary, Calgary, Alberta.
Beier, P. 1993. Determining minimum habitat areas and habitat corridors for cougars. Conservation Biology 7:94-108.
Craighead, F.L. 1994. Conservation genetics of grizzly bears. PhD Diss. Montana State University, Bozeman, Montana. 191 pp.
Craighead, F.L., and E. Vyse. 1996. Brown/grizzly bear metapopulations. In: D. McCullough (Ed.) Metapopulations and Wildlife Conservation Management. Island Press, Washington DC and Covelo California. Chapter 14: pp. 325-351.
Ehrlich, P. R. 1986. The Loss of Diversity. In: E. O. Wilson (Ed.) Biodiversity. National Academy Press, Washington D.C. pp. 21-27.
Gilpin, M.E., and M.E. Soulé. 1986. Minimum viable populations: Processes of species extinction. In: M.E. Soulé (Ed.) Conservation Biology: the science of scarcity and diversity. pp 19-34.
Greenwood, P.J. 1980. Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour. 28:1140-1162.
Harris, L. 1984. The Fragmented Forest: Island Biogeography Theory and the Preservation of Biotic Diversity. The University of Chicago Press, Chicago.
ICE6. 1994. ICE6 Tool Kit for Cumulative Effects Analysis. Systems for Environmental Management, Missoula MT.
Lovejoy, T. E. and others. 1986. Edge and other effects of isolation on Amazon forest fragments. In: Soule, M. E. (Ed.) Conservation Biology: The Science of Scarcity and Diversity. Sinauer and Associates, Sunderland, MA. pp. 251-285
McCord , C.M. 1974. Selection of winter habitat by bobcats on the Quabbin Reservation, Massachusetts. Journal of Mammalogy 55:428-437.
Noss, R.F., and L.D. Harris. 1986. Nodes, networks, and MUMs: preserving diversity at all scales. Environmental Management 10:299-309.
Noss, R.F. 1983. A regional landscape approach to maintain diversity. BioScience 33:700-706.
Noss, R.F. 1987. Protecting natural areas in fragmented landscapes. Natural Areas Journal 7:2-13.
Noss, R.F. 1992. The Wildlands Project: land conservation strategy. Wild Earth (special issue):10-25.
Noss, R.F., H.B. Quigley, M.G. Hornocker, T. Merrill, and P.C. Paquet. 1996. Conservation biology and carnivore conservation in the Rocky Mountains. Conservation Biology 10(4):949-963.
Paetkau D., L. Waits, P. Clarkson, L. Craighead, E. Vyse, R. Ward, and C. Strobeck. 1997a. Dramatic variation in genetic diversity across the range of North American brown bears. In Preparation.
Paquet, P., and A. Hackman. 1995. Large carnivore conservation in the Rocky Mountains: A long-term strategy for maintaining free-ranging and self-sustaining populations of carnivores. World Wildlife Fund Canada, Toronto, Canada.
Picton, H. 1986. A possible link between Yellowstone and Glacier
grizzly bear populations. Proceedings International Conference on
Bear Research and Management. 6:7-10.
Quigley, T.M., R.W. Haynes, and R.T. Graham (Eds.) 1996. Integrated Scientific Assessment for Ecosystem Management in the Interior Columbia Basin. USDA Forest Service General Technical Report PNW-GTR-382. Pacific Northwest Research Station, Portland, Oregon. pp 165-167.
Ruggiero, L.F., K.B. Aubrey, S.W. Buskirk, L.J. Lyon, and W.J. Zielinski (Eds.). 1994. The scientific basis for conserving forest carnivores: American marten, fisher, lynx, and wolverine, in the Western United States. USDA Forest Service General Technical Report RM-254. Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colorado. p. 133.
Saunders, D.A. and R.J. Hobbs (Eds.) Nature Conservation 2: the role of corridors. Beatty and Sons, Surrey UK. 442 pages.
Shaffer, M. 1981. Minimum population sizes for species conservation.. BioScience 31:131-134.
Shaffer, M. 1987. Minimum viable populations: Coping with uncertainty. In: M.E. Soulé (Ed.) Viable Populations for Conservation. Cambridge: Cambridge University press. pages 69-86.
Shaffer, M. 1978. Determining minimum viable population sizes: A case study of the grizzly bear (Ursus arctos L.). PhD Diss. Duke University.
Simberloff , D.S., J.A. Farr, J. Cox, and D.W. Mehiman. 1992. Consequences and costs of conservation corridors. Conservation Biology 1:63-71.
Simberloff , D.S., and L.G. Abele. 1976. Island biogeograhy theory and conservation practice. Science 191:285-286.
Soule, M.E., and M.E. Gilpin. 1991. The theory of wildlife corridor capability. In: D.A. Saunders and R.J. Hobbs (Eds.) Nature Conservation 2: the role of corridors. Beatty and Sons, Surrey UK. pp3-8.
USDA Forest Service. 1996. Status of the Interior Columbia Basin: summary of scientific findings. PNW-GTR-385. Portland, OR. 144pp.
USFWS. 1993. Grizzly bear recovery plan. U.S. Fish and Wildlife Service, Missoula, Montana. 181pp.
Walker, Richard
American Wildlands
40 E. Main St. #2
Bozeman MT 59715
tel: 406-586-8175
fax: 406-586-8242
rich@wildlands.org
Craighead, Frank Lance.
American Wildlands
40 E. Main St. #2
Bozeman MT 59715
406-586-8175
LCRAIGHEAD@AOL.COM