Max A. Moritz
David M. Stoms
Mark I. Borchert
Frank W. Davis
A Proposed Protocol for Identifying Potential Research Natural Areas with Gap Analysis Data
Abstract: In this paper we propose a protocol for identifying and evaluating candidate sites for the U. S. National Forest's Research Natural Area (RNA) program. The approach is explicit and repeatable and consists of the five following steps: 1) Vegetation Target Type Identification, 2) Quantification of Environmental Variation, 3) Calculation of Site Suitability, 4) Site Selection, and 5) Ground Truth. We also describe a prototype case study using this approach for Mixed Evergreen Forest in Los Padres National Forest in central coastal California. This case study demonstrates the usefulness of Gap Analysis data and GIS-based procedures in conjunction with applications outside GIS, such as vegetation classification and optimization programs. Results indicate that there are often complex trade-offs between efficiency (i.e, covering the most types with the least sites) and suitability (i.e., choosing the most ecologically appropriate sites) of solutions, even in a simple case such as the one described here.
INTRODUCTION
As humans continue to develop and urbanize formerly pristine wilderness areas, land preservation becomes an increasingly important issue. Preserving tracts of land maintains genetic and ecological diversity, while providing many other benefits such as reference areas for successional studies, control areas for manipulative research, and sites for educational activities. Decisions about how much land to designate and where nature reserves should be established often conflict with other interests. The number of protected sites and the number of variables to be evaluated in these decisions make the planning problem extremely complex. Therefore, conservation planning must effectively use existing data and sophisticated land allocation models to justify proposals for additional protected areas.
The USDA Forest Service's Research Natural Areas (RNA) program is one specific system of selecting and managing research sites. The RNA program was established in 1927 to identify, protect, and manage a representative sample of pristine areas of forest, shrubland, grassland, alpine, aquatic, and geologic types (U.S. Forest Service 1990). Forest Service guidelines recommend that new RNAs be sited in areas that best represent the ecological conditions needed to complete the system of natural areas and in areas where conflicting uses are minimal. Whenever possible, selected areas should show little or no evidence of major disturbances by humans (e.g., livestock grazing, timber cutting) for the past 50 years. Also, guidelines recommend the selection of entire small drainages of at least 121 ha (300 acres) in the western states.
The method for nominating candidate RNAs varies among vegetation types and locations. Site selection is relatively straightforward for types with limited distributions that provide few alternative sites. For more extensive vegetation types, site recommendations have been based on the knowledge of local or regional experts. Other sites have been nominated because they have few conflicts with other uses such as cattle grazing or timber harvesting, rather than because they were the best representatives of the types in the province. The current selection process has successfully identified some areas that contain occurrences of vegetation types noteworthy for their condition and structural and floristic properties. Nonetheless, this process could be strengthened by a more systematic and repeatable method for selecting alternative sites. In particular, it is difficult in the present approach to measure the degree to which a site typifies floristic and environmental properties of the vegetation type over the region.
Selecting new RNA areas is currently a somewhat ad hoc process that begins when a site is nominated by a specialist familiar with the vegetation type in the region. That site is then scrutinized on the ground by other scientists and, if approved, moved through additional review and administrative procedures. There is no formal basis for identifying a set of alternative sites and systematically choosing among them based on the degree to which they meet goals of representativeness, pristine condition, size, and configuration. It is generally not feasible to subject all occurrences of a type in a large region to detailed ground survey. We propose, however, that mapped information on vegetation, environmental factors, and land management can be used to narrow and prioritize the set of candidate sites to a few alternatives that can be analyzed in detail. Ideally, any map-guided procedure for initial screening and prioritization should be explicit, repeatable, and successful at locating a small set of the best candidate sites.
Because regional RNA systems are designed to represent multiple vegetation types, consideration should also be given to overall regional design. For instance, if good examples of different types co-occur in a watershed, it may be more efficient and practical to select that watershed for both types then to designate and manage separate watersheds for the different types. Also, effective monitoring of environmental change may depend on a well-designed network of RNA sites that efficiently capture floristic and environmental variation both within and among plant community types. This requires planning across types over the entire region, an application for which the Gap Analysis Program (GAP) was designed (Scott et al. 1993; Davis et al. 1995; Davis et al. 1996).
We have developed an approach for identifying and prioritizing sites for a regional RNA system for the U. S. Forest Service. This approach uses optimal siting techniques as part of a general process for RNA site selection based on a systematic description of vegetation and environmental variation in the region, in conjunction with analysis of patterns of vegetation, land ownership, and management (from the California Gap Analysis project). We envision this process as a preliminary step to precede more detailed ground survey and administrative review procedures as currently practiced.
Here we describe our approach to RNA screening and site prioritization and demonstrate its application for representation of Mixed Evergreen Forest types of Los Padres National Forest in central coastal California. This case study is described fully in Stoms et al. (in review), although more of the technical details will be discussed here. First we outline the proposed GIS-based procedure, followed by a description of specific methods and results of applying the approach to the Mixed Evergreen Forest type. GIS is used for identifying target types and their distribution with respect to land ownership, characterizing environmental variation of these types, determining suitability of the pool of candidate sites, and visualizing model results. The GAP database is the only source of regional data available for this type of application.
THE PROPOSED PROTOCOL
Our proposed protocol follows a sequence of five steps:
1. Vegetation Target Type Identification: Identify the subset of target vegetation types to be represented in RNAs (classification of GIS attribute data and GIS overlay);
2. Quantification of Environmental Variation: Characterize typical environment within the target types (GIS overlay and summary statistics);
3. Calculation of Site Suitability: Characterize the suitability of sites to represent each target type (GIS modeling);
4. Site Selection: Apply a siting model to identify the most efficient and/or suitable set of sites (linear programming model plus GIS for visualization);
5. Ground Truth: Evaluate the best sites in the field.
Vegetation Target Type Identification: The purpose of this step is to determine which vegetation types a particular national forest should be responsible for representing in an RNA. We suggest the types to be represented follow the proposed National Vegetation Classification System (NVCS; FGDC 1996) at the alliance level. Alliances are defined by their dominant canopy species within structural formations. The distribution of alliances on a national forest and in its ecoregion can be determined from the databases compiled as part of GAP (Scott et al. 1993). Although other selection criteria could be used, we propose target types be limited to those with at least half of their regional extent occurring on national forest system lands.
Quantification of Environmental Variation: Because areas are designated "in perpetuity" as RNAs, one must consider how likely it is that a candidate site will ensure the long-term persistence of a vegetation type in that location. By quantifying the amount of environmental variability exhibited for each type across a region, one gets an idea of the "typical" environmental characteristics for each type. It is well documented that the distributions of most plant species and communities exhibit unimodal patterns of frequency or abundance along environmental gradients (e.g., Whittaker 1975). More stable sites would be those near the center of their distributions (i.e., the mean or median values) in terms of environmental variability. Less stable sites would be unusual in terms of environmental variability (i.e., values in the tails of their distributions) and might be expected to show the greatest changes in plant species composition after an ecological disturbance such as fire. Quantifying variation in environmental characteristics and identifying typical environments can be accomplished by GIS overlay with simple summary statistics. In this context, candidate sites can be evaluated and down-weighted if they are considered atypical for the vegetation type in question.
Calculation of Site Suitability: This step relates the characteristics of target types to the pool of candidate sites. This relationship can be quantified as a suitability index which is calculated independently for each type at each site. Factors that determine suitability are a combination of the areal extent of the type at the site, plus the representativeness of the site for the target type in terms of the environmental variables. Both of these factors can be derived readily by GIS operations. If mapped data are available, past land uses such as logging and grazing history could also be used to rate sites or to eliminate heavily disturbed areas.
Site Selection: In this step, the "best" site or sites are identified that represent one or more of the target types for potential RNA designation. This selection can involve trade-offs between picking the most efficient set of sites (i.e., least number of sites with the most types present) and picking the most suitable sites (i.e., ecologically the most typical). As the number of target types, measures of suitability, and candidate sites increase, trade-offs between efficiency and suitability rapidly become difficult to evaluate without the aid of mathematical models.
Ground Truth: We do not presume that a GIS and site selection procedure will eliminate the need for field inventory of high priority sites. There will always be a need to validate GIS data on the ground, determine site condition, and collect site-specific data not available in a regional GIS database. Although this is critical in practice, we do not discuss it further in this paper.
LOS PADRES NATIONAL FOREST CASE STUDY
Overview
The Monterey Ranger District (MRD) of Los Padres National Forest covers approximately 125,000 ha in the rugged Santa Lucia Mountains. At various locations along the coast, mountains rise from sea level to elevations of more than 1500 m within a distance of 4 km and form a series of steep, closely set north-south trending mountain ridges and deeply dissected drainages which continue inland for 25 km before descending into the Salinas Valley.
Following the logic of the statewide RNA program, we assumed a regional perspective in defining and prioritizing types for representation in RNAs. Based on the biogeographic regionalization of California presented in Hickman (1993), the MRD falls entirely within the Central Western biogeographic region of California. The Central Western California region, occupying more than 37,000 km², is bounded by the Russian River on the north, the Santa Ynez mountains on the south, the Pacific Ocean on the west, and the lower boundary of the foothill woodlands in the San Joaquin Valley on the east.
Our analysis takes advantage of a recently completed GIS database for the region that provides relatively coarse but spatially comprehensive information on vegetation, land ownership and management. This database was prepared by the California Gap Analysis Project (Davis et al. 1995), which is part of the national GAP sponsored and coordinated by the Biological Resources Division of the U.S. Geological Survey (Scott et al. 1993). We have combined these data with digital information on environmental characteristics and followed the first four steps of the protocol described above to select candidate watersheds for the Mixed Evergreen Forest type in the MRD region:
1. Divide the Mixed Evergreen Forest type into sub-types that represent floristically and geographically distinct ecological entities (TWINSPAN classification of dominant vegetation species in GAP polygons) and assess ownership and management status of these sub-types to identify those predominantly on National Forest System lands (overlay vector boundaries and filter);
2. Determine the environmental affinities of each type (quantify variation in raster GIS layer representing elevation);
3. Quantify suitability of each candidate site for each type (calculate how far each site is from typical environmental conditions);
4. Apply a siting model to identify a set of watersheds that appear to be good candidates for representing the Mixed Evergreen Forest types (optimization model with ARCVIEW for visualization).
Vegetation Target Type Identification
The vegetation map and associated data on dominant overstory species were prepared for the California Gap Analysis Project using methods described by Davis et al. (1995). Vegetation of the Central Western region was delineated into 3,595 landscape units with an average size of 1,037 ha, and distributional information was provided for 162 dominant plant species, 88 community types and 15 land use/land cover types. Each map unit was described by up to three upland stand types and the riparian type considered the most widespread in the map unit. For each type, up to three dominant overstory species were listed, and this species list was used to assign the type to classes in existing vegetation or wildlife habitat classification systems.
Mixed Evergreen Forest is the dominant forest type at middle elevations where Mediterranean climate is not ameliorated by summer fog (Barbour and Major 1977). The Mixed Evergreen Forest type is so extensive and variable that it is unreasonable to represent the type in a single RNA. This forest type is composed of a mixture of two or more mostly broad-leaved, evergreen tree species such as Lithocarpus densiflorus, Arbutus menziesii, Quercus agrifolia, Q. chrysolepis, Q. wislizenii and Umbellularia californica. Winter deciduous species like Acer macrophyllum and Q. kelloggii may also be components of this forest along with some conifers, especially Pseudotsuga menziesii (Sawyer et al. 1977; Holland 1986). The California GAP database contains presence data for 13 tree and 4 shrub species in 425 mixed evergreen forest polygons of the Central Western region.
We subdivided this general type into floristically and geographically distinctive sub-types using two-way indicator species analysis (TWINSPAN; Hill 1979). This was accomplished by exporting GAP polygon species combinations to text files and creating matrices of presence/absence for each species in each polygon. In this way, polygons were classified as if they were individual vegetation plots, and significant species associations were identified for the region. Species with fewer than three occurrences in the data set were eliminated from the analysis. Based on the analysis we recognized twenty-three widespread sub-types.
Map units of the GIS vegetation coverage were then re-attributed according to the new Mixed Evergreen Forest sub-types and overlaid with a coverage of land ownership to determine the fraction of each sub-type represented on MRD. Rather than target all Mixed Evergreen Forest types in the region that occur on MRD, we set targets only for those sub-types with the majority (i.e., > 50%) of their distribution on the MRD, regardless of their management status in the region. Although this is in agreement with RNA goals of preserving a representative sample of typical vegetation types, future RNA siting exercises should investigate other cutoffs and additional vegetation sub-types.
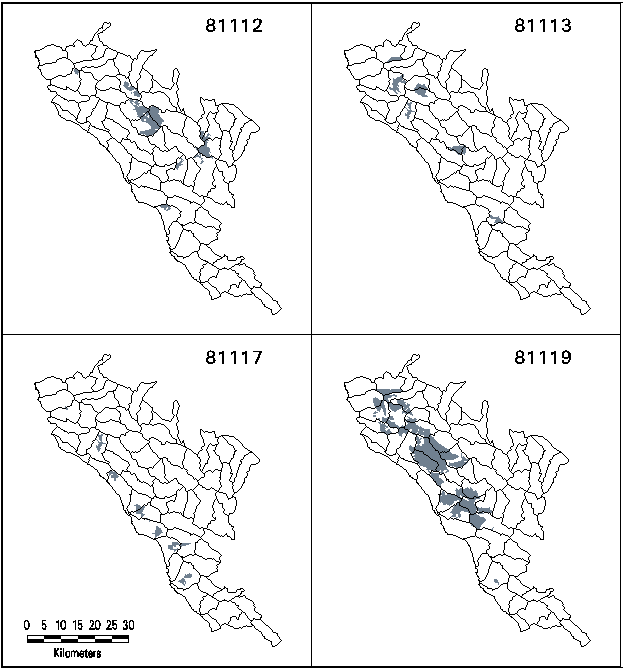
Of the 23 mixed evergreen forest sub-types identified in TWINSPAN analysis, four (type codes 81112, 81113, 81117, and 81119) have at least 50% of their mapped distribution within the region on the MRD (Table 1 and Figure 1). These four types were therefore selected as target types for representation in future RNAs.
Table 1. Percentage Ownership and Mapped Distribution of Mixed Evergreen Forest Target Types. Species codes are: ARME = Arbutus menziesii; LIDE = Lithocarpus densiflorus; QUAG = Quercus agrifolia; QUCH = Quercus chrysolepis.
| ClassCode | Dominant Species | LPNF % | Other Public % | Private % | Total Distribution (km²) | ||
| 81112 | QUAG | QUCH | -- | 75.5 | 2.4 | 22.1 | 53.9 |
| 81113 | QUAG | QUCH | ARME | 59.4 | 9.0 | 31.6 | 26.0 |
| 81117 | QUAG | ARME | LIDE | 51.3 | 17.6 | 31.1 | 32.0 |
| 81119 | ARME | QUCH | LIDE | 80.1 | 7.1 | 12.8 | 190.8 |
Figure 1. Mapped Distributions of Target Types. Solid lines are watershed boundaries in MRD region.
Quantification of Environmental Variation within Target Types
A modified measure for elevation was chosen as the environmental characteristic to determine whether a specific site was typical for a vegetation type for this case study. Variation due to changes in both elevation and latitude is captured through data representing "equivalent elevation" of an area, which is the actual elevation derived from a DEM that has been adjusted to account for distance from the equator. As such, equivalent elevation is useful only as a relative measurement to compare sites and is higher than actual elevations for central coastal California (e.g., Mixed Evergreen sites for this case study have an average equivalent elevation of approximately 3500 m). The latitudinal adjustment is 0.625 m of elevation per km of northness (Schoenherr 1992) and has been added to a DEM with a 100 m cell size. The ZONALSTATS command was used to calculate median equivalent elevation for each of the four mixed evergreen types (Figure 2) across the study area.
Figure 2. Boxplots of Variation in Equivalent Elevation for Target Types. The notched line in the center of the boxes indicates median values, and the size of the notch indicates approximate 5% confidence intervals about the medians. The size of boxes indicates the quartiles, and the outermost whiskers indicate 1.5 times the interquartile range; individual values outside the whiskers are shown as hollow circles.

Calculation of Site Suitability for Target Types
Based on the RNA Program's goal of selecting entire small drainages for RNA status, we adopted Calwater Planning Watersheds as the set of sites for our selection process. These watersheds, delineated by the California Department of Forestry and Fire Protection to support regional planning, average 2,750 ha in size in the study area. Although the planning watersheds used here are considerably larger than the RNA program's recommended minimum area of 121 ha, this scale of analysis is compatible with the resolution of available digital biological and environmental data and is consistent with our overall goal of prioritizing areas for more detailed analyses.
We used three factors to characterize watershed suitability for siting an RNA to represent Mixed Evergreen Forest types: 1) areal extent of all Mixed Evergreen Forest in the watershed, 2) the areal extent of the specifically targeted Mixed Evergreen Forest sub-types in the watershed, and 3) the degree to which environmental conditions in the watershed typify the conditions preferred by target sub-types in the region. These factors are described in more detail below.
We assume that an ideal RNA should not be an isolated island of a target community surrounded by distinctly different types. Accordingly, we computed the proportion of each planning watershed in all Mixed Evergreen Forest types combined, both target and non-target types. Watersheds with less than 15% coverage by Mixed Evergreen Forest and less than 300 ha of all target sub-types on were screened from the set of candidate sites.
Our second premise is that larger contiguous areas of a target type are more suitable for RNA designation than smaller areas, because they are likely to experience biophysical conditions and disturbance regimes more characteristic of that type. An area suitability index was computed for each vegetation polygon, being set to 0.0 if the area was less than 75 ha in a single vegetation polygon, while areas greater than 300 ha were assigned an index value of 1.0. All intermediate sizes were scaled linearly between 0.0 and 1.0.
Environmental suitability was calculated independently for each target type in each vegetation polygon as the absolute value of the difference between the regional median (equivalent) elevation for the type and the polygon's median elevation. This index was linearly scaled from the minimum to maximum values with 1.0 for the smallest difference and 0.0 for the largest difference (i.e., least typical elevation).
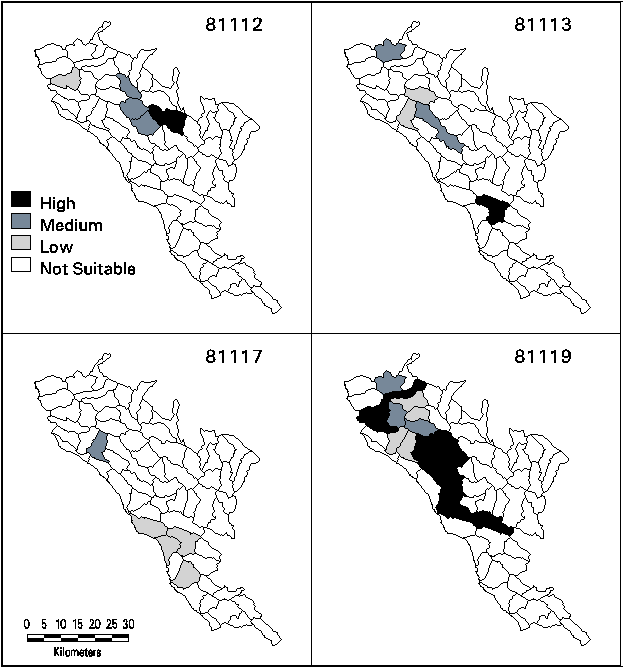
Three final steps were required to determine suitability of each remaining planning watershed from the individual polygon suitability scores. An overall suitability for each target type in each vegetation map unit was calculated simply as the product of the area suitability and environmental suitability, again giving potential values between 0.0 and 1.0. (The products were not rescaled from 0.0 to 1.0, however.) The maximum suitability index value for each type in each watershed was assigned as the site suitability for that type. The numerical suitability index was converted into Low, Medium, and High categories with breakpoints at 0.5 and 0.75.
After the screening step, 24 watersheds remained that could potentially represent one or more target types within at least 15% of their area in mixed evergreen landscape or at least 300 ha of one or more target types. Types 81112, 81113, and 81117 could each be represented by five sites (although usually not overlapping between types) and 81119 had 18 candidate sites. All four target types had High, Medium, and Low ratings in at least one watershed, except for 81117 which was not rated as High suitability in any watershed (Figure 3).
Figure 3. Mapped Suitability by Watershed for Target Types.
Site Selection
For this prototype application, with only four target types and 24 sites, the selection process could probably be achieved by a very simple modeling approach. However, for more complex problems with many sites and target types covering the full range of vegetation on a national forest, a computer algorithm similar to the one described here would be required. Even in this simple case, the trade-offs between efficiency (least number of sites) and suitability (the best sites) become difficult to evaluate, leading to the use of an optimization model.
The model is derived from a classic maximal covering location problem (MCLP) in which the most types are represented, or "covered", at a fixed number of sites (Church et al. 1996). The MCLP model uses standard operations research techniques to find an optimal solution, such that no more efficient solution can be found, although equally efficient solutions might exist. The original version described in Church et al. (1996) treated all sites with a target element as equally suitable, but here the basic MCLP model has been modified to accommodate variations in site suitability by user-specified weights. Thus, trade-offs between efficiency and suitability can be evaluated.
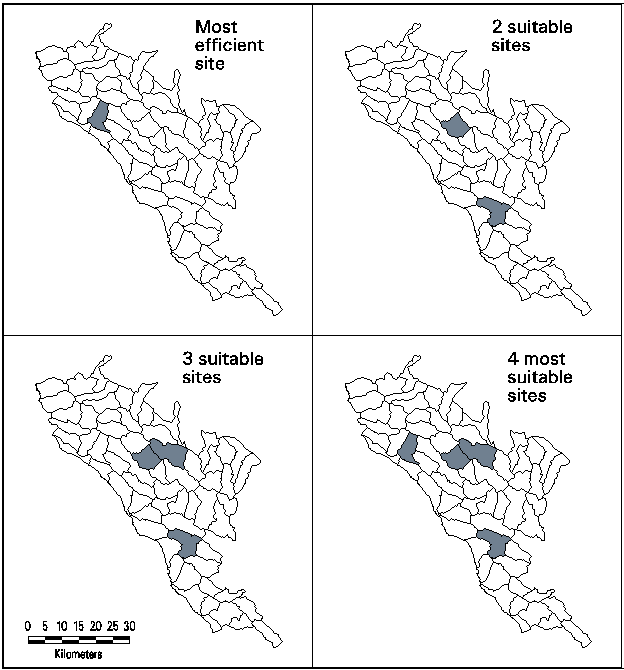
Several alternatives were run through the model using varying number of sites and weights on the suitability ranks. Here we present four alternatives to illustrate varying emphasis on efficiency versus suitability (Table 2 and Figure 4). Alternatives described here put increasing weight on higher suitability classes and allow more sites to be chosen in the final solution. The first alternative simply covers the most types in a single site, without regard to suitability (i.e., High, Medium and Low suitability classes are rated equally). Lion Creek represents the most "efficient" single site with three mixed evergreen types, but it is not necessarily the most suitable one. In fact, it rates no higher than Medium suitability for any type, and is rated Low for two types. When two sites can be selected and weights are increased for higher suitability, all four types can be represented with only one type (81117) covered with Low suitability. A three site solution with still higher suitability weighting increases the suitability of one of the types (from Medium to High for 81112) in comparison with the two site solution. The fourth alternative allows four sites, having more extreme weights on suitability to ensure the best environments for each type and putting no emphasis on efficiency. While selected sites do vary when suitability weights are changed, some watersheds tend to occur in many alternatives. For instance, Carrals Spring is the only site with High suitability for type 81113, and it is selected in all alternatives with more than one site and where High suitability is weighted more than Medium. Visualization of model results allows exploration of the trade-offs between the efficiency and suitability criteria as well as the number of selected sites.
Table 2. Watershed characteristics and results of alternative RNA selection model runs. Blanks in suitability ratings indicate the type did not occur in the watershed, at least not in adequate amount to be a representative of the type. "X" under the alternatives indicates watershed was selected in the alternative.
| Suitability Ratings | Alternative | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Watershed Name | 81112 | 81113 | 81117 | 81119 | 1 site | 2 sites | 3 sites | 4 sites | |
| Blackrock Creek | M | M | |||||||
| Pine Creek | H | ||||||||
| Danish Creek | L | ||||||||
| Upper Little Sur R. | L | H | |||||||
| Puerto Suello Ck. | M | ||||||||
| Miller Canyon | M | ||||||||
| Bruce Fork | L | ||||||||
| S. Fork Little Sur R. | H | ||||||||
| Blue Creek | L | M | |||||||
| Ventana Creek | L | ||||||||
| Up. Tassajara Ck. | M | H | |||||||
| Lion Creek | L | M | L | X | X | ||||
| N. Fork Big Sur R. | M | H | |||||||
| Rocky Creek | H | X | X | ||||||
| Low. Tassajara Ck. | M | H | X | X | X | ||||
| S. Fork Big Sur R. | H | ||||||||
| Higgins Creek | H | ||||||||
| Big Creek | H | ||||||||
| Big Devils Canyon | H | ||||||||
| Salsipuedes Creek | H | ||||||||
| Vicente Creek | L | ||||||||
| Carrals Spring | H | L | X | X | X | ||||
| Mill Creek | L | ||||||||
| Willow Creek | L | ||||||||
Figure 4. Map of Selected Sites for Four Alternative Solutions.
DISCUSSION AND CONCLUSIONS
The approach presented in this paper formalizes a GIS-based protocol for screening and prioritizing potential RNA sites that is less ad hoc and subjective than the process described in the Forest Service manual. The proposed protocol has two advantages over the current approach: 1) it considers the regional context in identifying target types to be represented by a national forest, and 2) it incorporates efficiency and suitability explicitly into the process. Efficiency is more important when the number of target types becomes large and sites can potentially represent more than one type. In the case study for Los Padres National Forest, with only four types and 24 sites, modeling was relatively simple and could almost be done by inspection. In a real situation with tens of types and hundreds of sites (e.g., Davis et al. 1996), conservation planning problems become so complex that computer algorithms become absolutely essential. Suitability is valuable in identifying sites that best represent the target types, and it should be incorporated into site evaluation. Here we only used median elevation to characterize environmental variation, but in the future we intend to consider more environmental variables (including fire regime) to obtain a richer set of descriptors. The fifth step in our proposed protocol, field validation and detailed inventory, will always be required, regardless of the quality of the GIS database, the sophistication of the analysis, or the ingenuity of the model.
Prioritizing potential RNA sites for detailed investigation should be performed in a regional context rather than with data restricted to lands managed by a single land steward or smaller geographic area. In many regions, GAP databases represent the first comprehensive source of information about natural plant communities and land ownership of adequate resolution to address this type of regional conservation planning. The nominal map scale of GAP databases is 1:100,000, with a minimum mapping unit of 100 ha for land cover types, and the classification attempts to match the NVCS alliance level. Thus GAP data are useful for intermediate scale inventories of vegetation types to be represented by an RNA system. They can help identify the set of target types that are best represented on National Forest System lands (or other land managing agency), characterize their physical environments, and relate those landscape level properties to a set of candidate sites such as planning watersheds. GAP data have also been used in siting alternative biodiversity management areas in the Sierra Nevada Ecosystem Project (Davis et al. 1996) and are currently being used for a similar planning effort for The Nature Conservancy in the Columbia Plateau ecoregion.
The case study presented in this paper used an optimization model to solve a weighted suitability maximal covering location problem. This model was external to the GIS, and data and results were passed between the two environments via ASCII text files. Gerrard et al. (in review) have recently adapted the ARC NETWORK module to present the basic the basic reserve siting problem to ArcInfo as a conventional facilities placement problem. This tighter integration of the model with the GIS is achieved through an innovative "logical" network, linking biodiversity elements to sites. Users of public-domain GAP data can then conduct their own explorations of potential reserve systems without having to acquire and master optimization software and reformat data for modeling or display. We believe it should be relatively straightforward to modify this NETWORK-based model to accommodate site suitability factors, further extending the utility of this approach.
ACKNOWLEDGMENTS
Support for this project was provided through a cooperative agreement from Los Padres National Forest. The California Gap Analysis Project was funded through grants from the National Biological Service, the National Fish and Wildlife Foundation, Southern California Edison Company, the Southern California Association of Governments, and the U. S. Forest Service. Computing facilities were provided through a grant from the IBM Corporation Environmental Research Program. We also gratefully acknowledge R.L. Church's help in running the optimization models.
REFERENCES
Barbour, M.G. and Major, J. 1977. Introduction. In: Barbour, M.G. and Major, J. (eds.) Terrestrial Vegetation of California, pp 3-11. California Native Plant Society, Sacramento.
Church, R. L., D. M. Stoms, and F. W. Davis, 1996. Reserve selection as a maximal covering location problem. Biological Conservation, 76: 105-112.
Davis, F. W., P. A. Stine, D. M. Stoms, M. I. Borchert, and A. D. Hollander. 1995. Gap analysis of the actual vegetation of California: 1. The southwestern region. Madroño, 42: 40-78.
Davis, F. W., D. M. Stoms, R. L. Church, W. J. Okin, and K. N. Johnson, 1996. Selecting biodiversity management areas. Pages 1503-1528 in Sierra Nevada Ecosystem Project: Final Report to Congress, vol. II, Assessments and scientific basis for management options. Davis: University of California, Centers for Water and Wildlands Resources.
Federal Geographic Data Committee, Vegetation Subcommittee. 1996. FGDC Vegetation Classification and Information Standards--June 3, 1996 Draft. FGDC Secretariat, Reston, VA, 35 p.
Gerrard, R., D. M. Stoms, R. L. Church, and F. W. Davis. Selecting conservation reserves using species covering models: Adapting the ArcInfo GIS. Submitted to Transactions in GIS.
Hickman, J. C., editor, 1993. The Jepson Manual: Higher Plants of California, University of California Press, Berkeley, California.
Hill, M. O. 1979. TWINSPAN, a FORTRAN program for arranging multivariate data in an ordered two-way table by classification of individuals and attributes. Ecology and Systematics, Ithaca, NY.
Holland, R. F., 1986. Preliminary descriptions of the terrestrial natural communities of California. State of California, The Resources Agency, Nongame Heritage Program, Dept. Fish & Game, Sacramento, Calif. 156 p.
Sawyer, J. O., Thornburgh, D. A. and Griffin, J. R. 1977. Mixed evergreen forest. In: Barbour, M.G. and Major, J. (eds.) Terrestrial Vegetation of California, pp. 359-382. California Native Plant Society, Sacramento.
Schoenherr, A.A. 1992. A Natural History of California. University of California Press, Berkeley, California.
Scott, J. M., F. Davis, B. Csuti, R. Noss, B. Butterfield, C. Groves, H. Anderson, S. Caicco, F. D'Erchia, T. C. Edwards, Jr., J. Ulliman, and R. G. Wright. 1993. Gap analysis: A geographic approach to protection of biological diversity. Wildlife Monographs, 123: 1-41.
Stoms, D. M., M. I. Borchert, M. A. Moritz, F. W. Davis, and R. L. Church. A spatial analytic approach to siting Research Natural Areas. Submitted to Natural Areas Journal.
USDA, Forest Service. 1990. Forest Service Manual, FSM 4000-Research, WO Amendment 4000-90-1. Washington, D. C.
Whittaker, R. H. 1975. Communities and Ecosystems. 2nd ed. Macmillan, New York, NY.
Max A. Moritz
Department of Geography
University of California
Santa Barbara, CA 93106-4060, USA
Phone: 805-893-7815
FAX: 805-893-3146
Email: maxm@geog.ucsb.edu
David M. Stoms
Institute for Computational Earth System Science, University of California, Santa Barbara, CA 93106-3060
Mark I. Borchert
Los Padres National Forest, Goleta CA 93117
Frank W. Davis
Department of Geography, University of California, Santa Barbara, CA 93106-4060, USA