Jonathan Proctor
Michael Beltz
William Haskins
Four habitat variables were analyzed in relation to prairie dog colony locations on the Charles M. Russell and UL Bend National Wildlife Refuges using an ArcInfo Geographic Information System (GIS). A classification tree and a logistic regression statistical program searched for patterns between prairie dog presence and: 1) vegetation, 2) slope, 3) soil texture, and 4) soil depth. The dataset consisted of a complete census of the study site - 488,695 pixels of 30m x 30m, each coded with the above information. Both tests found vegetation and slope to correlate well with prairie dog presence. Soil texture correlated only minimally, and soil depth did not appear to be a significant factor.
A model was developed with six habitat categories based on the classification tree results (which split the data into categories based on the probability of prairie dog presence within each combination of variables) and was applied to the study site. The model was then extrapolated to south Phillips County, Montana. All known prairie dog town locations (mapped between 1979 and 1997) were overlaid and corresponded well with these categories. Most towns centered on the preferred habitat category, with presence in less suitable categories occurring primarily in relation to these towns.
Management implications for the study site and south Phillips County are discussed, with special consideration given to identifying core prairie dog habitat areas and their relation to the future of the prairie dog ecosystem and the ongoing black-footed ferret reintroduction program.
The black-tailed prairie dog (Cynomys ludovicianus) is a large, colonial, burrowing rodent of the squirrel family (Sciuridae) found on the short- and mixed-grass prairies of the Great Plains region of North America. At one time the prairie dog may have been the most abundant mammal in the region (Koford 1958), possibly numbering as many as 5 billion individuals (Seton 1929). Black-tailed prairie dogs historically occupied a significant portion of the Great Plains, estimated between a minimum of 2.8 percent to 10 to 20 percent of the region (Flath and Clark 1986, Summers and Linder 1978). Estimates of total area occupied range from 100 million acres to 700 million acres (Knowles and Knowles 1994, Seton 1929, Cully 1989).
The black-tailed prairie dog is distinguished from the three species of white-tailed prairie dogs by its geographic range (the others do not occur on the Great Plains), its more colonial nature, and its reddish-brown fur and black-tipped tail. It occurs at elevations ranging from 915 to 1,830 meters and digs extensive burrow systems with large mounds 15-20 cm high. Black-tailed prairie dog densities average 15/ha, with a range of 5-33/ha (Fagerstone and Ramey 1996).
Prairie dogs maintain complex species interactions by creating and providing important or essential habitat conditions (e.g., food, shelter) for several wildlife species of the Great Plains ecosystem. This includes species that are now in jeopardy, such as the black-footed ferret (endangered), mountain plover (candidate species), ferruginous hawk (sensitive), swift fox (candidate species), and western burrowing owl (sensitive) (Knowles and Knowles 1994; BLF and Sharps 1994). Many of these declines have been attributed to the decline of the prairie dog.
Prairie dogs also change their surrounding environment. For example, they alter vegetative processes by maintaining vegetation in an early growth stage, decreasing vegetative height, increasing bare ground, and increasing the percentage of forb cover (Koford 1958). This provides a diversity of habitat on the plains essential to wildlife species that depend on these conditions. Prairie dogs also alter long-term soil-building processes through bioturbation, or mixing of soil horizons. This in turn leads to new soil types (Thorp 1949; Koford 1958).
Bison, pronghorn, and cattle prefer grazing on prairie dog colonies because of the greater nutritional value per unit biomass of the vegetation found here (Coppock et al. 1983), while prairie dogs rely on these ungulates to reduce vegetation height in tallgrass regions, where prairie dogs cannot maintain shortgrass habitat alone (Sharps and Uresk 1990). In general, species richness appears significantly higher in prairie dog colonies than in the surrounding landscape (Reading 1993; BLF and Sharps 1994).
Due to their vast number and extent, prairie dogs must have greatly affected the structure and function of the Great Plains region. Despite such importance, humans have historically placed a negative value on prairie dogs, and have been lagely responsible for reducing the area occupied by prairie dogs by an estimated 90 percent (Flath and Clark 1986) to 98 percent or more (Miller et al. 1994) throughout North America. In Eastern Montana, the prairie dog currently occupies an estimated 0.17 percent of the landscape (Knowles 1995). These reductions are due to habitat destruction, poisoning, sport shooting, and the recent spread of sylvatic plague.
Public land agencies-including the U.S. Bureau of Land Management, U.S. Forest Service, and state land and wildlife management agencies-continue to manage prairie dog populations at numbers which are a fraction of historic levels (USFS 1986; USBLM 1992). Although prairie dogs still number a few million in isolated pockets scattered across much of their historic range, this severe reduction has essentially removed the disturbance function of the prairie dog on the grasslands, and numerous species that require such disturbances have subsequently plummeted in numbers.
If we wish to ensure the long-term viability of the entire prairie dog ecosystem, we must identify and protect the remaining habitat and locate unoccupied potential habitat in which restoration efforts may occur. Protection of the remaining 1-2% of fragmented prairie dog towns alone may not be enough to maintain the entire prairie dog ecosystem, including its disturbance function. For example, too few prairie dog complexes have been identified to date to ensure the successful reintroduction of the black-footed ferret (Reading et al., 1997).
This study provides a methodology for creating habitat maps outlining suitable BTPD habitat on the Charles M. Russell and UL Bend National Wildlife Refuges (CMR) and similar lands. These maps may be used to identify areas where successful augmentation efforts or natural colonization are most likely to succeed, as well as prioritizing areas in which to seek conservation easements on private lands for the protection and restoration of the prairie dog ecosystem.
A procedure for determining suitable BTPD habitat is also needed to implement the Montana Prairie Dog Management Guidelines. These guidelines call for site-specific management plans that describe the occupied and potential ranges of prairie dogs in the planning area. A detailed map is also suggested. Mapping methodologies to rapidly assess habitat suitability are, therefore, key to successful implementation of these guidelines.
This study proposes a method of outlining suitable prairie dog habitat on a wide scale with a minimum of cost and effort. By combining existing vegetative, slope, and soil data with a Geographic Information System (GIS), prairie dog habitat maps outlining varying degrees of suitability can be created for large areas. Such maps are more coarse in scale than detailed site-specific maps that depend on extensive site-specific measurements (i.e., vegetation height), but maps at this scale may prove invaluable for identifying wildlife corridors, core reserves, and isolated colonies. This information may be used to outline critical wildlife habitat and/or develop plague management plans. Also, public land agency wildlife budgets may prohibit analysis at greater levels of detail.
In order to delineate suitable BTPD habitat as a subset of the total landscape, first it must be shown that BTPDs selectively "choose" from the resources available to them. Several studies imply such resource selection. For example, Clippinger (1989) developed a habitat suitability index model for BTPDs and Tepley et al. (1990) used this information to produce a GIS model of potential and preferred prairie dog habitat, but the variables on which these studies are based remain untested. Reading (1993) did test a list of variables with a GIS and found BTPD occupancy on smaller slopes, one soil association (Elloam soils), and BLM ownership to be significantly greater than expected. He suggests analyzing vegetation, shooting impacts, proximity to other colonies, and associations with heavy livestock use and adding this to his data to create a predictive model of prairie dog colony expansion.
This study utilizes Reading's (1993) slope factor, drops the ownership factor (as not pertinent to the goals of this thesis), and alters the soil factor to allow its use across areas with various soil associations. It also includes Reading's suggestion of a vegetation factor analysis, but does not add the other suggestions for the following reasons: 1) vegetation information was created from satellite imagery for all of eastern Montana. The vegetation classifications inherently include all factors that impact vegetation enough to alter its biomass or species composition, including the impacts from livestock grazing or prairie dog occupancy. This is especially true given that the data is so recent and vegetation has remained relatively constant in this area over the past several decades; 2) shooting does not occur within the Refuge; and 3) adding proximity to other colonies as a factor would have significantly complicated the process, and the importance of this factor can be inferred from the final maps.
The Charles M. Russell and UL Bend National Wildlife Refuges (hereinafter referred to as CMR) encompass 1,094,301 acres of land and water, 760,000 acres of which are federal lands. Within the refuge's boundary are also Fort Peck Reservoir, state lands, and private lands. The study site consists of approximately 236,233 acres of land (369 square miles) within the CMR. This area lies in the western-most portion of the CMR including those areas of the refuge that fall within the following 7 1/2 minute topographic quadrangles: Grand Island, Bell Ridge West, Bell Ridge East, Sagebrush Reservoir, Blizzard Reservoir, Lake Reservoir, Karsten Coulee, Pea Ridge, Mitchell Crossing, Hessler Ridge, Carter Coulee, Kepple Bottom, Hanson Flat, Dry Coulee, Chain Buttes, and Locke Ranch. Excluded from study within these areas are those lands known to be unsuitable habitat (i.e., forested lands, water, steep slopes). This leaves 148,766 acres (233 square miles) which were included in the study.
Three main landforms dominate the study site: uplands, breaks, and floodplains. Elevation ranges from 2,000 feet above sea level to 3,200 feet. The Missouri River bisects the refuge and study site, carving 500- to 1000-foot-deep valleys. Floodplains have been submerged by Fort Peck Lake through all but the western edge of the refuge (this area is included in the study site). Uplands consist of rolling prairies dissected by intermittent streams. Breaks lie adjacent to the Missouri River in a band 2-10 miles wide, and make up approximately 40-50 percent of the land within the CMR (USFWS 1985). The CMR receives 12-13 inches of precipitation per year, about 70 percent occurring from April-September. Runoff often exceeds 50 percent due to the heavy-textured soils. Temperatures range from an average low in January of 0 degrees Fahrenheit to average highs in the 80s in summer. Lightning storms in late summer often result in wildfires. Soil moisture is rapidly lost in summer due to high temperatures, low humidity, and regular winds (USFWS 1985).
Besides numerous general descriptions from early travelers of the plains, virtually no historic prairie dog data exists from which a "natural" prairie dog ecosystem can be described. The Charles M. Russell and UL Bend National Wildlife Refuges (CMR) best approximate prairie dog habitat in eastern Montana because: 1) prairie dogs have been relatively free from human control efforts since 1964, longer than any other area of eastern Montana (Knowles 1982). As a result, they have been able to expand to occupy what is thought to be a large percentage of their suitable habitat; 2) this area is part of a relatively large and biologically important prairie dog complex (Reading et al., 1997); and 3) accurate prairie dog distribution data is available.
The following research hypotheses were examined: black-tailed prairie dogs (BTPDs) choose: 1) short- to medium grassland cover types more than expected; 2) slopes of 0-8% more than expected; 3) soils ranging in texture from clay to sandy loam more than expected; and 4) soils with depths greater than 60 inches more than expected.
Results were used to identify and map habitat categories on the study site. The model was then extrapolated to adjacent south Phillips County, Montana to create a second map of prairie dog habitat categories across a much greater area.
Vegetation, slope, and soil are considered to be the factors affecting BTPD resource selection. The research hypothesis consists of the subsets of each factor which BTPDs are presumed to prefer. These factors and cutoff levels were selected after a thorough literature review of related studies, expert interviews, and spot checks of several black-tailed prairie dog colony locations (see "sources of variation" section below for a thorough defense of these assumptions). Factors were also chosen for their ease of collection (e.g., vegetation height does not need to be measured in the field) and for their applicability throughout the region.
Each factor was divided into several subsets as follows, with the research hypothesis subsets in bold print:
All areas classified as urban, agricultural, forestlands, water, riparian, or alpine in the vegetation category were eliminated from consideration, as were all areas with slopes greater than 25%. This was done because in general, prairie dogs do not inhabit these areas (Hall 1981), although they may on occasion inhabit undeveloped areas within urban areas or agricultural areas that have been abandoned. Also, by removing these unlikely categories from consideration, the remaining possibilities would be more accurately defined as being suitable or not.
Soil was characterized by three attributes: depth, texture, and drainage class. These soil attributes were chosen because they are believed to be the factors important to burrow construction (Osborn 1942, Koford 1958), and because they can be applied to other regions regardless of the specific soil types because all soil classifications contain these individual factors. Drainage class was dropped from the final analysis due to the fact that all soils within the study site were classified as well drained.
Vegetation: BTPDs inhabit short- and mixed-grass prairies in the semi-arid plains (Clippinger 1984; Reid 1954), and are able to spread into tallgrass prairie following heavy grazing by ungulates (Osborn 1942; Schaffner 1926). Vegetation height in BTPD colonies ranges from 7 to 13 cm (Agnew et al. 1986) and up to 64 cm (Clark et al. 1982). This vegetation height is necessary for visibility which allows protection from predators (Hoogland 1981). In short- and mixed-grass prairie, BTPDs alone are able to maintain this vegetation height. However, in tallgrass prairie, BTPDs rely on ungulates to reduce vegetation height; if ungulates are absent, BTPD colonies will be reduced in size and eventually eradicated (Osborn and Allen 1949). A rare BTPD town has been found in a Cottonwood stand or shinnery savanna, but this is only on the edge of large towns when severe overgrazing has occurred (Reid 1954; Osborn 1942). Sagebrush is not a complete barrier to BTPD dispersal, as they can progressively invade and cut these plants (Reid 1954), although they are still dependent on livestock to graze any tallgrasses in the area (Osborn 1942). BTPDs seem to prefer disturbed areas in grasslands (Koford 1958; Knowles 1982). Old fields are especially attractive to BTPD habitation (Reid 1954), and BTPDs thrive in overgrazed areas (Koford 1958).
Grassland vegetation is also essential for food requirements. Stomach exams of BTPDs in Montana have found 98.6% vegetative content (Kelso 1939). Stomach and fecal exams of BTPDs in South Dakota found five major grasses: western wheatgrass, blue grama, buffalo grass, sixweeks fescue, and tumblegrass (Wydeven and Dahlgren 1982). These species and others (including hairy grama, hairy triodia grass, and sand dropseed) have been identified in several studies as species utilized by BTPDs (Knowles 1982; Clippinger 1984). All of these species are found in short- and mid-grass prairie (MT GIS lab 1995).
Slope: BTPDs prefer flat areas or gentle slopes. Several studies have measured slopes on prairie dog towns. The findings are listed in the following table:

One additional study found that BTPD colonies are located on flatter terrain than are randomly located polygons (Reading 1993).
Soil: Cover may be the most important requirement for BTPDs; soil provides this requirement. BTPDs require well-drained soils that are capable of retaining water for burrow stability. BTPDs occur in soil textures ranging from clays to sandy loams (Proctor 1995; Reid 1954; Sheets 1970; Knowles 1982). Sandy soils are avoided (Osborn 1942; Reid 1954; Knowles 1982). BTPDs are attracted to disturbed soils, such as livestock watering sites and old homesteads (Knowles 1982).
Burrow construction requires soil of sufficient depth. Sheets (1970) excavated 18 burrows and found their depths to range from 3 to 14 feet (7' mean, 8.5' median). Only 3 burrows were less than 60 inches. Also, soils with depths less than 5 feet are classified as poorly drained. However, BTPDs have been observed to burrow through soft bedrock such as shale (Knowles 1982).
Soil information was derived from Natural Resource Conservation Service (NRCS) data. The soil survey for Phillips County has yet to be published. A GIS soil map of Phillips county (north study site) was created from scanned images provided by NRCS which were cleaned up by removing dangling nodes and connecting missing segments using PC ARC/INFO version 3.4.2 (Esri 1994b). A GIS soil map of Fergus and Petroleum Counties (south study site) was created by photographing the published NRCS paper surveys, scanning the negatives with a Polaroid slide scanner, and then digitizing the scanned images with Arcview 3.0a for Microsoft Windows (Esri 1994a).
Each soil polygon was then coded by three categories: depth, texture, and drainage class, and each of these categories was divided into the classes listed in Table 2.1. The polygons were then converted to 30 x 30 meter pixels to match the vegetative data, and a separate layer was created for both soil depth and for soil texture.
Vegetative data for eastern Montana was provided by the Montana Wildlife Spatial Analysis Lab. The Lab classified the vegetation in the scene which covers the study area from satellite imagery taken in 1991. A vegetation code key was created for this purpose. Each pixel was classified within this key based on its reflective properties.
All pixels labeled with vegetation categories which are not capable of being inhabited by prairie dogs were dropped from this analysis (i.e., forested areas, water). Of the categories that were left, 23 fall within the study site (6 grassland categories, 7 shrubland categories, 3 shrub-grassland complex categories, and 7 barren land categories).
Slope data was derived from 30 x 30 meter USGS Digital Elevation Models (DEMs). This information was condensed into the categories listed in Table 2.1. These categories were chosen because they match NRCS soil information and because differences at smaller slopes may be more crucial for predicting suitable habitat than differences at larger slopes. All areas with slopes over 25% were dropped from analysis because in general these slopes fall outside of the range of prairie dogs.
Prairie dog locations on the CMR-as well as on neighboring BLM lands and the Fort Belknap Indian Reservation-have been accurately located and mapped recently using a Global Positioning System (GPS). Surveys done before GPS became available were originally mapped by hand on 7.5 minute topographic maps, but have since been digitized from the mapped locations. CMR prairie dog town locations were mapped in 1979, 1984, 1988, and 1995 by CMR employees. These maps were combined to create a map of maximum-known occupied prairie dog area. This combined data map was then used for the CMR study site map. The south Phillips County map combined these prairie dog town locations with prairie dog town locations for the entire region mapped between 1979 and 1997.
The CMR study site was divided by the Missouri River into two sections for analysis. The prairie dog populations south of the river may not have recovered fully from the days of poisoning (Knowles, pers. comm. 1998), and thus the results of this area may not be as revealing as those north of the river. The south data was used only to check inferences made from the north dataset. The north study area included 488,695 pixels of data, and the south study area contained 180,520 pixels of data.
An ArcInfo 7.1.1 for Microsoft Windows NT (Esri 1997) was used to create a separate map layer for each factor. The scale for all maps is 1:24,000, and the minimum mapping unit (MMU) for all maps is 30 x 30 meters. Maps are in NRIS format (Albers projection, in meters).
Layers representing soil depth, soil texture, slope class, vegetation class, and prairie dog presence/absence were then overlaid, excluding all pixels known to be unsuitable for prairie dog occupancy, such as cliffs, forests, and water. A dataset for each remaining 30 x 30 meter pixel was then created. The number of pixels included in this analysis totaled 669,215, which equals an area of 956 square kilometers (369 square miles), or 95,641 hectares (236,233 acres). All pixels with the same combination of these five variables were then grouped and tallied. The north dataset contained 517 unique combinations, and the south dataset contained 316.
Two steps of data analysis occurred. First, a classification tree (S-Plus version 3.4, StatSci, 1996) was computed for the north and south datasets using all available variables (vegetation, slope, soil texture, soil depth, and prairie dog presence/absence). A classification tree is an exploratory tool used in this case to find which variables seem to be most strongly associated with prairie dog presence. Because classification trees are known to over-fit the data, cross-validation was used to estimate how large a model was needed (the statistical program automatically withholds a portion of the information and cross-validates the results against this subset).
Second, a logistic regression model (S-Plus version 3.4, StatSci,
1996) further analyzed the datasets to explain the variation between
available and occupied habitat. A new vegetation-related classification
with 5 categories was created based on biomass to facilitate analysis.
This reclassification was done in the following manner: the vegetation
information for each pixel contained not only a vegetation category
but also a value based on the Modified Normalized Differenced
Vegetation Index (MNDVI). This commonly-used value-a ratio between
the red band and near infrared band-correlates well with biomass
(Nemani et al. 1993). Therefore, 5 MNDVI categories were delineated
based on the breaks in the MNDVI values that were used to classify
grassland vegetation categories in the original development of
the vegetation code key (Table 2.3). These categories were then
applied to all vegetation types. 
Each pixel was reclassified based on its MNDVI value, and a new dataset was created by replacing the vegetation code column with the biomass column. In this analysis, 188 unique combinations occurred in the north dataset and 147 occurred in the south dataset.
Summary tables of prairie dog presence as a function of the categories show that prairie dogs in the study site are associated with certain vegetation types, MNDVI values, slopes, soil depths, and soil textures. Presence is measured as the proportion of each category occupied by prairie dogs (i.e., in Table 3.1, 51.7% of the north dataset pixels with vegetation code 3130 contain prairie dogs). Both vegetation and MNDVI tables are listed because each was used in the statistical tests: the classification tree used the vegetation data and logistic regression used the MNDVI data.
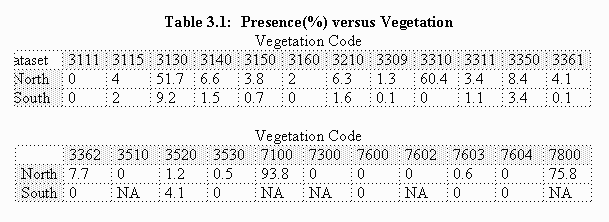


Classification tree analysis revealed a strong pattern between vegetation category and slope with prairie dog presence/absence. A weaker association with soil texture was also noted. No association was noted with soil depth. Plots of predictive deviance against model size suggest that trees with no more than six leaves are needed or should be used for these datasets, so further splits were avoided. If an association does exist with soil depth, it occurs beyond this cutoff level.
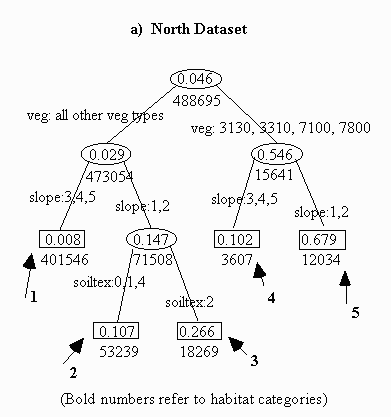
For the north dataset, the classification tree found that BTPDs choose: (1) four specific vegetative categories more than expected. The classification tree separated vegetation codes 3130 (very low cover grasslands), 3310 (Salt-Desert shrub), 7100 (dry salt-flats), and 7800 (mixed barren sites) from the other 19 categories. This suggests that prairie dogs are associated with these four vegetation types;
(2) slopes of 0-4% more than expected. The classification tree separated slopes of 0-4% from slopes of 4-25%, suggesting that slopes greater than 4% are not a significant factor associated with prairie dog presence; and
(3) clay-loam soils more than expected. Only clay-loam soils were separated as a significant factor of prairie dog presence, and only then in cases of less-desired vegetation types and small slopes.
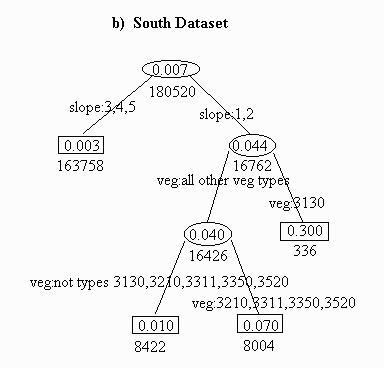
The south dataset contains less information due to the lower percentage of prairie dog towns (0.7% versus 4.5% in the north). The first split for the south separated slopes 1 and 2 from the steeper slopes, and the second split separated vegetative type 3130 from the others (no types 7100 or 7800 were observed, and all 452 sites with vegetation code 3310 had an absence of prairie dogs).
Table 3.4 shows a preference for soil depths greater than 60" (category 4), but this preference was not strong enough for the classification tree to separate within the chosen level of confidence.
Logistic regression results using MNDVI were similar to the classification tree model in that slope was the single most important variable with MNDVI being second and soil texture (in the north dataset only) coming in third. For the north dataset, slope accounted for 32% of the overall variation, and MNDVI accounted for 19% of the variation (for the south dataset, the amounts are 20.5% and 6% respectively).
To interpret logistic regression coefficients, the following formula is needed: Probability of Presence = exp (a + bi + cj) 1 + exp (a + bi + cj) where a (the intercept) is given as 0.7175; bi is 0 for MNDVI=1, -0.9449 for MNDVI=2, -1.889 for MNDVI=3, -2.557 for MNDVI=4, and -6.07 for MNDVI=5; and cj is 0 for slope=1, -1.133 for slope=2, -3.108 for slope=3, -6.743 for slope=4, and -8.692 for slope=5.
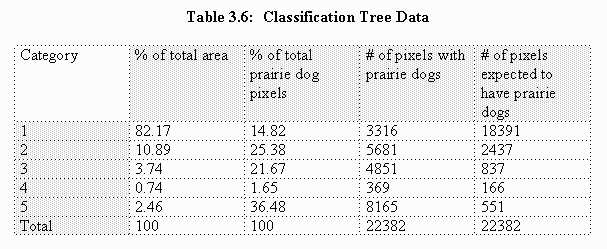
The north dataset classification tree habitat category model (Figure 3.1) was applied to the study site to create a habitat map outlining six habitat suitability categories, the sixth habitat category being areas excluded from the study (Map 1). Category 5 contains prairie dogs on 67.9% of pixels and represents the preferred habitat. With a 26.6% occupancy rate, category 3 represents potential prairie dog habitat. Categories 2 and 4 have almost identical occupancy rates at 10.7% and 10.2% respectively, also representing potential habitat. With only 0.8% occupancy, category 1 represents unsuitable habitat. Areas excluded from the study contained no prairie dogs and are therefore also unsuitable.
Within the study site, 23,335 acres fall within categories 2-5. This equals about 9.9% of the total area. In comparison, only 2,756 acres, or 1.2% of the study site, falls within the preferred habitat category 5. If these categories are considered potential prairie dog habitat, then the north dataset, with a 4.6% prairie dog occupancy rate, had prairie dogs on 46% of potential habitat prior to the recent plague-related decline.
The model was also applied to south Phillips County (Map 2). Only one vegetation category (7601-shrub badlands) occurred in south Phillips County that did not occur in the study site and represents a minute percentage of the total area (well less than one percent). These areas are labeled "unclassified" .
This new map necessitated removing the split separating categories 2 and 3 because the soil texture variable that separates these categories is not available for Phillips County (Figure 3.2). The colors between the study site map and the south Phillips County map correlate, although pink now represents both the pink and yellow categories from the CMR study site map.
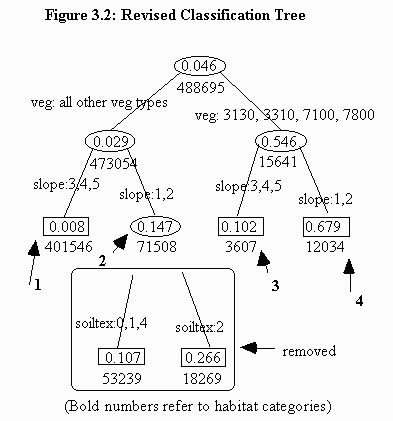
Total potential habitat within south Phillips County equals 1,137,853 acres. Of this total, 143,748 acres fall within the preferred category 4. The maximum extent of prairie dogs between 1979 and 1997 totaled only 34,255 acres, or 3% of potential habitat.
The categories in which prairie dogs were located were similar to the findings in the CMR study site. The largest single percentage of all prairie dogs occurred in category 2 (a combination of two categories in theCMR study site). Second was preferred category 4 (Table 3.8).
Prior studies found a high correlation between prairie dog presence and certain soil associations (Knowles 1982, Reading and Matchett 1997). However, when the individual soil factors were studied in conjunction with slope and vegetation factors, soil depth was not found to be a factor while soil texture was only a minor factor. It is likely that the results from prior studies are due to correlations that these soil associations have with small slopes and certain vegetation types.
For the vegetation factor, statistical tests analyzed both the MNDVI value (biomass) and vegetation code. Vegetation code is of greater use in defining areas suitable for prairie dogs than MNDVI because prairie dogs are known to be associated with areas of low biomass (MNDVI correlates with biomass), but whether this is a factor which they select in colonizing an area or whether this is a result of their presence is not clear. It could be in fact that prairie dogs prefer areas with greater biomass, but their presence over time results in the low biomass values associated with these towns. A correlation with specific vegetation categories, however, would aid in identifying suitable habitat regardless of its exact biomass at any specific point in time.
Having stated this preference for using vegetation type over MNDVI, it should be noted that one common theme between the four vegetative codes that correlate well with prairie dog presence-3130 (very low cover grasslands), 3310 (Salt-Desert shrub), 7100 (dry salt-flats), and 7800 (mixed barren sites)-is their low biomass. Clearly, prairie dogs select areas with relatively low biomass.
Because results between the classification tree test and the logistic regression test were similar and validated each other, and because of the greater simplicity of the classification tree-which is easier to interpret and use than logistic regression coefficients-the classification tree model was used to define prairie dog habitat. The classification tree is also preferable to the logistic regression model because the former can be used for either vegetation code or MNDVI value, while the latter can only interpret MNDVI value.
These maps may be used to predict where future expansion of BTPD colonies is most likely to occur, either independently or through reintroduction efforts. They may also aid in comprehending the current situation by outlining how much suitable habitat a certain area contains, and the percentage of this suitable habitat that is currently occupied. They also outline areas where management efforts should be concentrated for the greatest benefit.
CMR map: The vast majority of prairie dog towns are centered on or at least occur partially within category 5 (preferred vegetation and 0-4% slopes), which strongly suggests the importance of this category as preferred prairie dog habitat (this category also contains the largest single percentage of prairie dogs). When pixels with prairie dog presence fall within categories 2,3, or 4, they often occur at the edges of towns that center on category 5. These categories (2,3, and 4) therefore appear to be suitable habitat to varying degrees and primarily as a result of their association with category 5.
With a 26.6% occupancy rate, category 3 (secondary vegetation, 0-4% slopes, and clay-loam soils) appears to be a secondary category of preference. Category 2 (secondary vegetation, 0-4% slopes, and other soil textures), however, contains a larger number of prairie dog pixels than category 3 because it covers a larger area, even though it has a lower occupancy rate (10.7%).
With a 10.2% occupancy rate, category 4 (preferred vegetation and 4-25% slopes) is of limited importance here due to the small area it represents. The limiting factor for this category may be that vegetation codes 3100, 3310, 7100, and 7800 rarely occur on slopes greater than 4%.
With only a 0.8% occupancy rate, category 1 is of little value to prairie dog habitat except where it borders existing towns within the other categories. Even though this category covers the majority of the study area (82%), only two small prairie dog towns occur solely within category 1. These are located along the banks of the Missouri River, possibly attributable to a factor not considered (assumed to be due to concentrated human impacts, which seem to attract prairie dogs).
Finally, no prairie dog towns occur within the areas excluded from study . This appears to justify the assumptions made in rejecting these areas as suitable habitat.
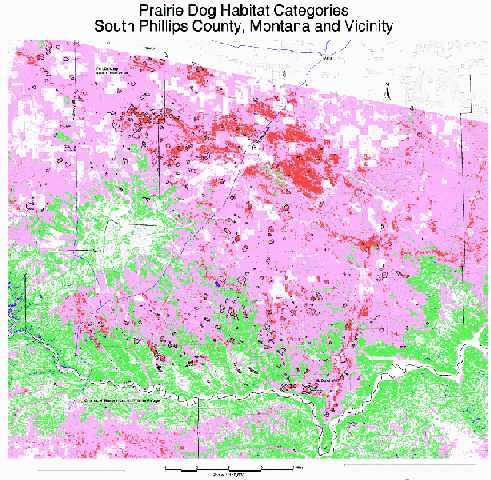
South Phillips County Map: When the model was extrapolated to the neighboring region, the same patterns occurred. Most towns centered on the preferred category (in this case, renumbered as 4) and in several cases stopped at this category's boundaries. Also, a minimal percentage of the unsuitable category 1 contained prairie dogs. In this map, however, the majority of the area is considered potential habitat, as opposed to the study site (included in this larger map) in which the majority of the area was unsuitable. This shows the relative value of the region as prairie dog habitat. The geographic patterns of preferred habitat (denoted as red on this map) are easily located within the region.
Curiously, a large area of preferred habitat contains relatively few prairie dog towns (located due south of Malta and southeast of the "U.S. Highway 191" label on the map). This area is primarily private land, however, and may be poisoned on a more regular basis than public land. A much greater extent of prairie dog towns (both in number and in size) occurs on the same habitat type to the immediate west of this area on the Fort Belknap Indian Reservation, possibly due to differences in poisoning programs.
This map shows clear patterns of core areas of preferred prairie dog habitat and potential connecting corridors amidst a larger pattern of semi-potential and unsuitable habitat. This information could be used to prioritize management of certain areas to benefit the larger prairie dog ecosystem.
The results of this study are most accurately applied to the Northern Great Plains shortgrass prairie ecoregion. Factors change as one moves out of this region (e.g., slope appears to be more restrictive for prairie dog occupancy further south). The results do not reveal directly why certain factors are selected or avoided, but only tabulates the amount to which these factors are associated with existing prairie dog towns within the study site.
This model may be applied across all of eastern Montana once the vegetation data for this area is finished. A prairie dog habitat suitability map may then be produced with at least the five categories now available, and all six categories in areas where NRCS soil surveys have been completed. For application in these areas, however, vegetation should be similar to the 23 codes found within the study site. If a significant difference in vegetation occurs, this model may be of more limited value; it will still identify areas where prairie dogs are likely to succeed, but it may miss other suitable lands based on factors not considered in the study site.
Similar tests should be conducted in relatively natural prairie dog ecosystems in other ecoregions to create more accurate models for these regions. Few areas exist; Theodore Roosevelt National Park in North Dakota and Badlands National Park in South Dakota are two possibilities.
This model for predicting potential prairie dog habitat may be used to develop and/or improve prairie dog ecosystem management plans for lands in eastern Montana. Below are some examples, beginning with the study site itself.
The CMR developed a set of goals and objectives based on the laws, orders, and policies that guide its management (Executive Order 7509 and the National Wildlife Refuge System Administration Act of 1966). Wildlife objective 8 is to "Maintain viable prairie dog towns totaling no less than 5,000 acres and no more than 10,000 acres on suitable areas with sizes and patterns desirable for black-footed ferrets. Minimize conflicts with adjacent landowners" (USFWS 1985). The study site map outlines these suitable areas and identifies areas where future prairie dog colonies are most likely. The maps also show where suitable habitat exists near private land. These areas may be managed to prevent prairie dog colonization by, for example, removing livestock grazing which leads to shorter vegetation height and thereby increases the potential for prairie dog colonization.
Wildlife objective 2 is to "Maintain habitat for and reintroduce a minimum of six pairs of black-footed ferrets on six or more prairie dog towns when animals are available," and objective 11 is to "Reintroduce...swift fox into suitable habitat" (USFWS 1985). Ferrets are now being reintroduced and need more prairie dog towns to ensure a viable future. Swift fox may be reintroduced in the future if/when ferret reintroduction succeeds. Because swift fox densities are highest in areas with extensive prairie dog towns, prairie dog maps may also aid in this effort. Finally, because prairie dog towns in this area are important habitat for mountain plovers (a candidate species), prairie dog habitat maps may help define the potential of the CMR for mountain plover habitat.
Beginning in the summer of 1997, the CMR and private individuals began relocating prairie dogs onto plagued-out towns in an attempt to reestablish these areas for the benefit of the black-footed ferret reintroduction program and for mountain plover recovery. The study site map may help to identify appropriate translocation areas, especially areas that have not been occupied by prairie dogs in the recent past.
The prairie dog habitat map shows that, prior to plague in 1992, much of the primary habitat was occupied. Comparing habitat suitability on the CMR to neighboring areas in south Phillips County, it is clear that the CMR contains relatively little habitat, and the two main habitat areas that do exist are geographically separated from each other. For improved management of the black-footed ferret recovery program, the BLM lands with much greater suitability should play a greater role in this effort.
The Judith Valley Phillips Resource Management Plan states that "BLM, in cooperation with the FWS and MDFWP, would maintain the existing prairie dog habitat and distribution on BLM land within the 7 km Complex based on a 1988 survey" (USBLM 1992). In 1988, BLM lands within the 7 km Complex contained 12,346 acres of prairie dog towns. Between 1991 and 1996, sylvatic plague ran through the area and wiped out 70% of the prairie dog complex. The BLM has initiated a voluntary shooting ban on BLM lands, but the number of prairie dogs in the area remains far below plan objectives. An action plan to address how the BLM will return prairie dogs to 1988 levels will be developed in the first half of 1998, and the habitat map of south Phillips County may aid in developing a successful plan.
The prairie dog habitat map may be used to identify the best locations within south Phillips County in which to encourage or reestablish prairie dogs to create such corridors. Areas should be identified which would connect towns crucial to black-footed ferret recovery and reduce conflicts with adjacent private landowners.
The CMR and BLM may also use these maps to identify locations in which to establish or reestablish prairie dog towns to connect the ferret reintroduction site in the UL Bend area with the Manning Corral prairie dog town (recently wiped out by plague).
Although plague is not fully understood, it is thought that a diverse pattern of connected colonies of varying sizes as well as isolated colonies of various sizes is the best condition to ensure the future of prairie dogs and associated species. This model may identify isolated towns, and locate suitable areas in which to promote new isolated towns which may survive future plague epidemics.
Other areas of significant prairie dog habitat on public lands exist in eastern Montana, such as the Tongue River Valley. But the agencies which manage these lands (Custer National Forest and BLM's Powder River Resource Area) do not have such plans.
Conservation biologists have promoted such strategies for conserving numerous wildlife species and habitat. Prairie dog habitat may be outlined with this model and used as the basis for a conservation strategy as proposed in general terms by Noss and Cooperrider (1994) and specifically to the prairie dog ecosystem by Wuerthner (1997). Such a strategy is needed to ensure the long-term viability of the entire prairie dog ecosystem including the several dependent species in jeopardy as well as the overall ecosystem functions provided by significant numbers of prairie dogs.
When the vegetation map of eastern Montana is completed, the model can be used to identify significant areas of preferred habitat on public lands throughout the state. These areas could then form the "core reserves" necessary for such a proposal.
An absolute minimum of 10 such core reserves must be identified nation-wide in order to attain the goals of the black-footed ferret recovery program (USFWS 1988). Development of these core reserves should also consider other species' needs. Because this study site and its larger prairie dog colony complex has been identified as nationally significant for prairie dogs, black-footed ferrets, and mountain plovers (Knowles 1995, Reading 1993, Olson and Edge 1985), this area should constitute one such core reserve.

Agnew, W., D.W. Uresk, and R.M. Hansen. 1986. Flora and fauna associated with prairie dog colonies and adjacent ungrazed mixed-grass prairie in western South Dakota. J. Range Manage. 39(2):135-139.
Anderson, E., S.C. Forrest, T.W. Clark, and L. Richardson. 1986. Paleobiology, biogeography, and systematics of the black-footed ferret, Mustela nigripes(Audubon and Bachman), 1851. In The black-footed ferret. Great Basin Naturalist Memoirs. 8:11-62.
Biodiversity Legal Foundation and J.C. Sharps. 1994. Petition to classify the black-tailed prairie dog (Cynomys ludovicianus) as a category 2 candidate species pursuant to the administrative procedures act and the intent of the endangered species act. Letter to the U.S. Fish and Wildlife Service dated Oct. 21. 82 pp.
Clark, J.W., T.M. Campbell III, D.G. Socha, and D.E. Casey. 1982. Prairie dog colony attributes and associated vertebrate species. Great Basin Nat. 42(4):572-582.
Clippinger, N.W. 1984. Habitat suitability for the black-tailed prairie dog at Rocky Mountain Arsenal. U. of Colorado, B.A. thesis. 94 pp.
Clippinger, N.W. 1989. Habitat suitability index models: black-tailed prairie dog. U.S. Fish and Wildlife Service. Biological Report 82(10.156) 21pp.
Coppock, D.L., J.E. Ellis, J.K. Detling, and M.I. Dyer. 1983. Plant herbivore interactions in a North American mixed-grass prairie. Oecologia 56:10-15.
Cully, J.F., Jr. 1989. Plague and the prairie dog ecosystem: importance for black-footed ferret management. In The prairie dog ecosystem: managing for biological diversity. Clark, T.W., D.K. Hinckley, and T. Rich (eds.). Montana BLM Wildl. Tech. Bull. No. 2.
Dalsted, K.J., S. Sather-Blair, B.K. Worcester, and R. Klukas. 1981. Application of remote sensing to prairie dog management. J. Range Manage. 34(3):218-223.
Environmental Systems Research Institute, Inc. 1994a. ArcView. Version 3.0a for Microsoft Windows. Redlands, CA.
Environmental Systems Research Institute, Inc. 1994b. PC ARC/INFO. Version 3.4.2. Redlands, CA.
Environmental Systems Research Institute, Inc. 1997. Understanding GIS: the ArcInfo method. Version 7.1.1 for Microsoft Windows. Redlands, CA.
Fagerstone, K.A. and C.A. Ramey. 1996. Rodents and lagomorphs. In: P.R. Krausman, ed. Rangeland Wildlife. Denver: The Society of Range Management: 83-132.
Flath, D.L. and T.W. Clark. 1986. Historic status of black-footed ferret habitat in Montana. Great Basin Nat. Mem. 8: 63-71.
Hall, E.R. 1981. The mammals of North America, 2nd ed. New York: J. Wiley and Sons. 1181pp.
Hoogland, J.L. 1981. The evolution of coloniality in white-tailed and black-tailed prairie dogs. Ecology 62: 252-272.
Kelso, L.H. 1939. Food habits of prairie dogs. U.S. Dept. Agric., Cir. 529, 15 pp.
Knowles, C.J. 1982. Habitat affinity, populations, and control of black-tailed prairie dogs on the Charles M. Russell National Wildlife Refuge. U. of Montana, Dept. of zoology. Ph.D. thesis. 171 pp.
Knowles, C.J. 1995. A summary of black-tailed prairie dog abundance and distribution on the central and northern great plains. Faunawest Wildlife Consultants, Boulder, MT.
Knowles, C.J. 1998. Personal communication.
Knowles, C.J. and P.R. Knowles. 1994. A review of black-tailed prairie dog literature in relation to rangelands administered by the Custer National Forest. USDA Custer National Forest, Billings, MT. 61 pp.
Koford, C.B. 1958. Prairie dogs, whitefaces, and blue grama. Wildl. Monogr. 3: 1-78.
Manly, B.F., L.L. McDonald, and D.L. Thomas. 1993. Resource selection by animals: statistical design and analysis for field studies. Chapman and Hall, London. 177pp.
Miller, B., G. Ceballos, and R. Reading. 1994. The prairie dog and biotic diversity. Cons. Biol. 8: 677-681.
Montana Black-footed Ferret Working Group. 1988. Montana prairie dog management guidelines. Bureau of Land Management, Billings. 14pp.
Montana GIS Lab. 1995. Custer National Forest (eastern Montana, Dakotas) vegetation and land cover classification system modified from Montana and Idaho vegetation and land cover classification system of December 1995. Unpub. U. of Montana, Missoula.
Nemani, R., L. Pierce, and S. Running. 1993. Forest ecosystem processes at the watershed scale: sensitivity to remotely-sensed Leaf Area Index estimates. Int. J. Remote Sensing 14: 2519-2534.
Olson, S.L. and D. Edge. 1985. Nest site selection by mountain plovers in northcentral Montana. J. Range Manage. 38: 278-280.
Osborn, B. 1942. Prairie dogs in shinnery (oak scrub) savannah. Ecology 23: 110-115.
Osborn, B. and P.F. Allan. 1949. Vegetation of an abandoned prairie dog town in tall grass prairie. Ecology 30: 322-332.
Proctor, J.D. 1995. A case for change in prairie dog management on Montana's state lands. Unpub. U. of Montana, Missoula. 20pp.
Reading, R.P. 1993. Towards an endangered species reintroduction paradigm: a case study of the black-footed ferret. PhD. dissertation, Yale University, 552 pp.
Reading, R.P., T.W. Clark, A. Vargas, L.R. Hanebury, B.J. Miller, D.E. Biggins, and P.E Marinari. 1997. Black-footed ferret (Mustela nigripes): Conservation update. Small Carnivore Conservation 17: 1-6.
Reading, R.P. and R. Matchett. 1997. Attributes of black-tailed prairie dog colonies in northcentral Montana. J. Wildl. manage. 61(3); 664-673.
Reid, N.J. 1954. The distribution of the black-tailed prairie dog in the Badlands of southwestern North Dakota. State Univ. of Iowa, Iowa City. 30pp.
Schaffner, J.H. 1926. Observations on the grasslands of the central United States. The Ohio State University Press. Columbus, OH. 56pp.
Seton, E.T. 1929. Lives of game animals. Doubleday, Dovan, and Co. Inc. Garden City, N.Y.
Sharps, J.C. and D.W. Uresk. 1990. Ecological review of black-tailed prairie dogs and associated species in western South Dakota. Great Basin Naturalist Memoirs 50: 339-345.
Sheets, R.G. 1970. Ecology of the black-footed ferret and the black-tailed prairie dog. M.S. thesis. South Dakota State University, Brookings. 33pp.
S-Plus version 3.4, StatSci, Seattle, WA, 1996
Summers, C.A. and R.L. Linder. 1978. Food habits of the black-tailed prairie dog in western South Dakota. J. Range Manage. 31: 134-136.
Tepley, B., F. D'Erchia, and T. Schulz. 1990. Sighting of black-tailed prairie dog towns as potential reintroduction sites for the black-footed ferret. Colorado State U., unpublished. 22pp.
Thorp, J. 1949. Effect of certain animals that live in soils. Sci. Monthly, 68: 180-191.
Tileston, J.V. and R.R. Lechleitner. 1966. Some comparisons of the black-tailed and white-tailed prairie dogs in north-central Colorado. Amer. Midl. Nat. 75:292-316.
U.S. Bureau of Land Management. 1992. Judith Valley Phillips resource management plan and final environmental impact statement. Lewistown, MT. 436 pp.
U.S. Fish and Wildlife Service. 1985. Final environmental impact statement for the management of Charles M. Russell National Wildlife Refuge. Denver, CO. 453 pp.
U.S. Fish and Wildlife Service. 1988. Black-footed ferret recovery plan. Denver, CO. 154 pp.
U.S. Forest Service. 1986. Custer National Forest plan. Billings, MT.
Wuerthner, G. 1997. Viewpoint: The black-tailed prairie dog-headed for extinction? J. Range Manage. 50:459-466.
Wydeven, P.R. and R.B. Dahlgren. 1982. A comparison of prairie dog stomach contents and feces using a microhistological technique. J. Wildl. Manage. 46(4):1104-1108.
Jonathan Proctor
Predator Project
Box 6733
Bozeman, MT 59771
406-587-3389
jonathan@wildrockies.org
Michael Beltz
The Ecology Center, Inc
801 Sherwood, Suite B
Missoula, MT 59802
406-728-5733
beltz@wildrockies.org
William Haskins
The Ecology Center, Inc
801 Sherwood, Suite B
Missoula, MT 59802
406-728-5733
haskins@wildrockies.org