Calculating Landscape Fragmentation using a Gradient-based Approach
David M. Theobald1 and N.T. Hobbs2
1Natural Resource Ecology Lab, Colorado State University, Fort Collins, CO 80523-1499
2
Habitat Section, Colorado Division of Wildlife, Fort Collins, CO 80524(Paper presented at the 1999 Esri User Conference, San Diego, CA)
Abstract
Many ideas in landscape ecology extend from the concept of patches imbedded in a matrix. The patch/matrix concept is limited because it is insensitive to variation among species in their responses to landscape structure. Moreover, this insensitivity makes it very difficult to understand differences in patch and matrix quality. Such differences are critical to predicting effects of natural and anthropogenic disturbance. We offer an approach that uses allometric scaling of animal movement requirements at three time scales to create patch maps reflecting inter-specific differences in responses to landscape structure. Levels of organization are individual, population, and meta-population. We assign values to map cells based on the amount of habitat within an area, defined by requirements for home range and minimum population density.
Introduction
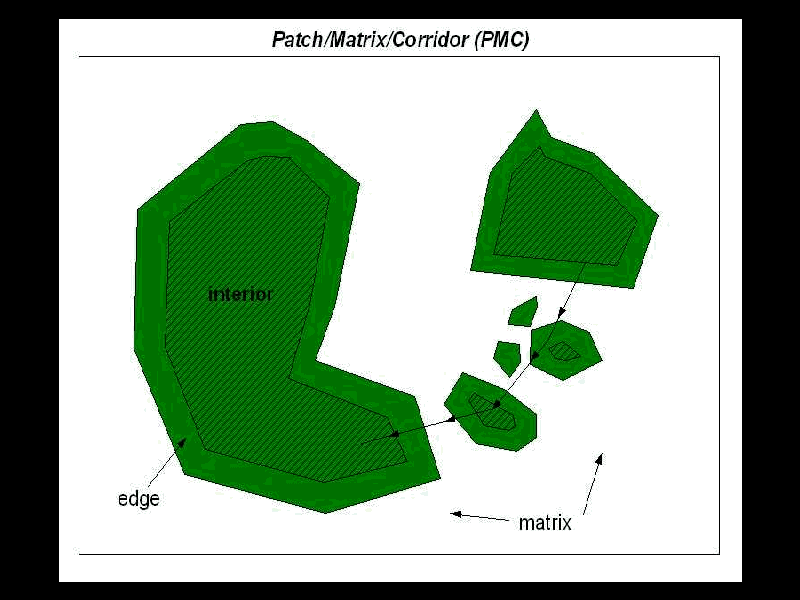
A mainstream approach to landscape ecology is based on the patch/matrix (PM) conceptualization of landscapes (Forman and Godron 1986) (Figure 1). This approach is rooted in island biogeography theory (MacArthur and Wilson 1967), where patches are considered islands and the matrix is represented as an inhospitable "sea" (Wiens 1994; Wiens 1996). The approach is straightforward and accessible because it corresponds to human scales of perception of the landscape and it easily integrates with traditional cartographic representations of landscapes using categorical maps (Gustafson 1998). Furthermore, analysis of these landscape maps can be accomplished easily through existing geographical information systems (GIS) tools (e.g., Haines-Young et al. 1993). As a result, the PM approach has been popular with practicing landscape ecologists, especially those in the planning field (e.g., Dramstad et al. 1996; Collinge 1996).
However, an important challenge for the field of landscape ecology is to move beyond simply providing general principles, yet be useful in practical management situations (Wiens 1999). We argue here that the PM approach is limited because it cannot easily incorporate functional responses of species to landscape structure. Rather, we advocate more fully incorporating functional response into landscape analysis, to develop an approach to studying landscapes that "embraces complexity rather than imposes simplicity on them" (Wiens 1999).
We find a number of situations in which detailed understanding of biological mechanisms that operate at the landscape scale are overlooked in the typical PM approach, but which need to be explicitly incorporated into a broader framework for understanding landscape structure. For example, the basis for most landscape ecology studies is a vegetation map. Patches of vegetation are commonly represented by polygons in a categorical map created either through photo-interpretation of aerial photography or aggregation of neighboring cells from a classified remotely-sensed image.
A number of aspects of these vegetation patch maps, though generally recognized, are usually overlooked. Representing patches as discrete entities (e.g., by a polygon) ignores both the fuzziness of the boundary and within-patch heterogeneity (Gustafson 1998). Non-contiguous patches of vegetation can be functionally integrated if a species or process of interest operates at a scale that can span patches (With and Crist 1995). Finally, uncertainties associated with vegetation data seldom are reflected in derived patch maps (Flather et al. 1997) even though all maps have a certain level of accuracy (Goodchild and Gopal 1986) and there are well-established methods of error assessment in land cover mapping (e.g. Congalton 1991).
Typically, suitable habitat near the edge of a patch is lost due to the "edge effect" as a result of various processes, such as increased predation and differences in environmental characteristics. The depth of the "edge effect" is typically modeled using a uniform distance (e.g., Reed et al. 1996), even though the processes that create edge are known to vary considerably, for example by aspect and edge contrast (Chen et al. 1995). Again, methods have been developed to vary the edge distance (e.g., Baskent and Jordan 1995) but are seldom employed in landscape analysis.
Patches are typically considered to be imbedded in a matrix of an inhospitable (non-habitat) intervening landscape. This matrix is generally considered to be biologically inert and, as a result, isolation of patches is generally considered simply in terms of Euclidean distance between patches (e.g., Keitt et al. 1997; Schumaker 1996). However, the matrix is not "ecologically neutral" (Wiens et al. 1993) and species’ movement through the landscape is affected not only by their probability of encountering patch edges, but how they perceive and respond to heterogeneity within the matrix. Matrix properties such as edge contrast, vegetation structure, and land use influence species movement.
We also seek to find an approach that serves as "middle-ground" between simple computation of landscape metrics (O’Neill et al. 1988) and current "state-of-the-art" metapopulation modeling (McCullough 1996). The application of metapopulation theory to understanding the consequences of landscape pattern change is limited because it requires a number of conditions that are not commonly met and are difficult to demonstrate. That is, metapopulation theory requires that local populations occur in discrete patches yet are within range of colonization, that no single population is large enough to persist alone, and population dynamics of different patches are asynchronous. Finally, it cannot provide insight into situations where local extinction occurs as a result of removal of a patch from land use change (Wiens 1996).
We assert that until approaches are developed that make the behavioral response to landscape structure explicit it will remain difficult to demonstrate linkages between landscape structure and ecological processes and to understand the consequences of land use change on the population viability and distribution. In this paper we develop an approach to define patches at three levels of organization: individual, population, and meta-population. We use allometic scaling of relationships to parameterize our models. After developing this methodology, we illustrate the differences between this gradient-based approach and the typical PM approach to landscape characterization. Importantly, this approach offers a way to screen potentially imperiled species by identifying critical thresholds and characteristics landscape patterns that result from known or predicted landscape changes.
Methodology
Our primary goal is to develop a methodology to define landscape structure that incorporates the way species respond to a landscape. We identify patches at three levels of organization: individual, population, and meta-population. One of the challenges of improving models to incorporate biological mechanisms is an increased demand on model parameterization. We use allometric relationships to parameterize our approach (Gaston and Blackburn 1996; Peters 1987).
Individual
At the fine-scale, movement is characterized by foraging, when species are maximizing their energetic return through foraging behavior (Hobbs in press). Species "integrate" resources over a local area and so definition of patches must reflect this scale of behavior (Addicott et al. 1987). The typical approach to identify patches at this point is to aggregate all adjacent (4 or 8-neighbors) that are contiguous. However, as discussed above, the patches that result are simply an artifact of the scale of the map and not based on how species (or processes) respond to landscape structure, and therefore landscape indices derived from these maps, such as number of patches, are unreliable.
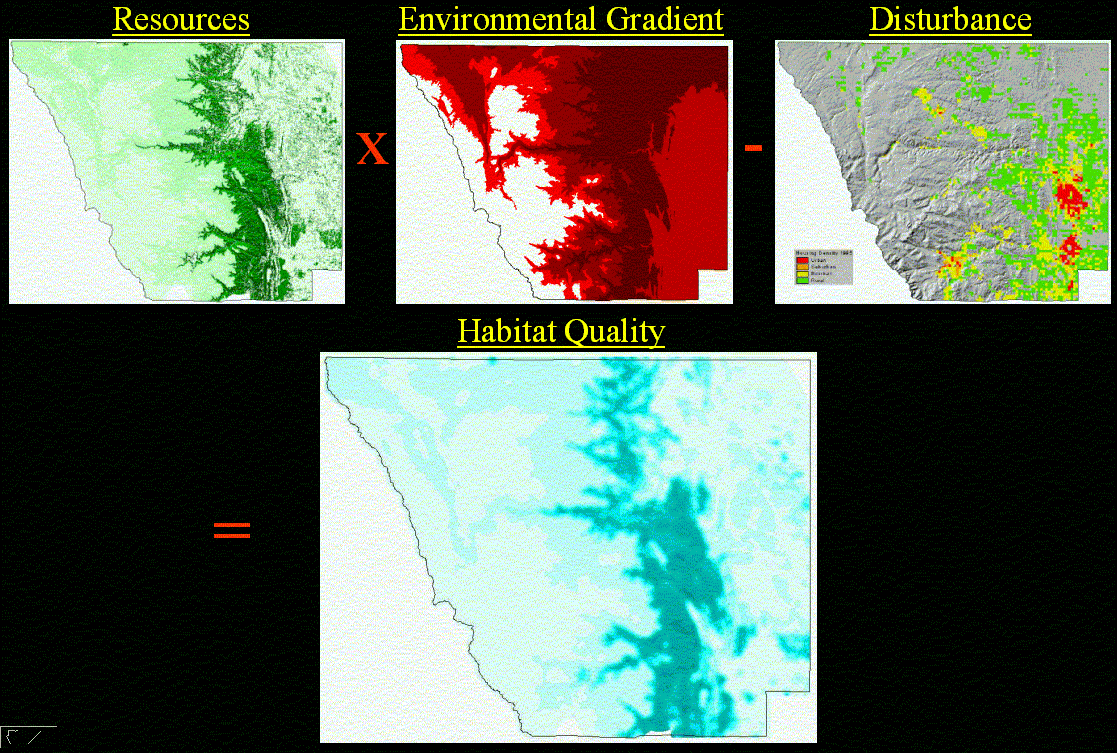
First we define resources, most commonly defined in terms of vegetation or land cover types, that serve as habitat for a given species. Typically this is done through a species-vegetation affinity table which contains the vegetation types utilized by a species (e.g., Ciacco et al. 1995; Edwards et al. 1996; White et al. 1997). We extend this approach by allowing the affinity value to range from 0.0 to 1.0, denoting the strength of the association. If an error matrix has been built for the vegetation image, uncertainty can be incorporated by multiplying the probability that a location mapped as a particular type is correct by the affinity score. A more elaborate approach would be to incorporate errors of omission and commission for species that utilize more than one vegetation type through the use of fuzzy logic (Shenk et al. 1997). This approach computes the degree of membership in each vegetation type for each location (pixel) on a map. Frequently other environmental gradients, such as elevation, slope/aspect, precipitation, and solar exposure, are used to modify the distribution of potentially suitable habitat for a species. Again, this is typically represented by a binary map, but we extend the values to range from 0.0 to 1.0.
The quality of vegetation in many locations is modified by either in-situ or nearby natural or human-caused disturbance. Of particular concern is the need to incorporate impacts of human-caused disturbances on habitat quality, typically from land use conversion. This requires a relationship to be developed between the disturbance (e.g., road or housing density) and the impact on habitat use (e.g., Theobald et al. 1997). Typically, the "edge effect" reduces patch quality near the edge, but we extend our approach to allow both lowered and increased habitat quality as a function of distance to edge. For interior species, habitat quality near the patch edge can be lowered, but for edge species, patch core areas may be poorer habitat. Furthermore, we allow the edge effect distance to change based on the adjacent patch type.
The habitat quality (Q) at a particular point (e.g., a pixel) is a function of four components (Figure 2):
Q = f(R,S,D,E) (1)
where resources, environmental factors, disturbances, and edge effect are denoted by R, S, D, and E, respectively.
We define patches at the individual level based on home range requirements. The area of home range (I) is based on an allometric relationship to body size (B) (Equation 2) (Harestad and Bunnell 1979):
I = 1.166M1.06 (2)
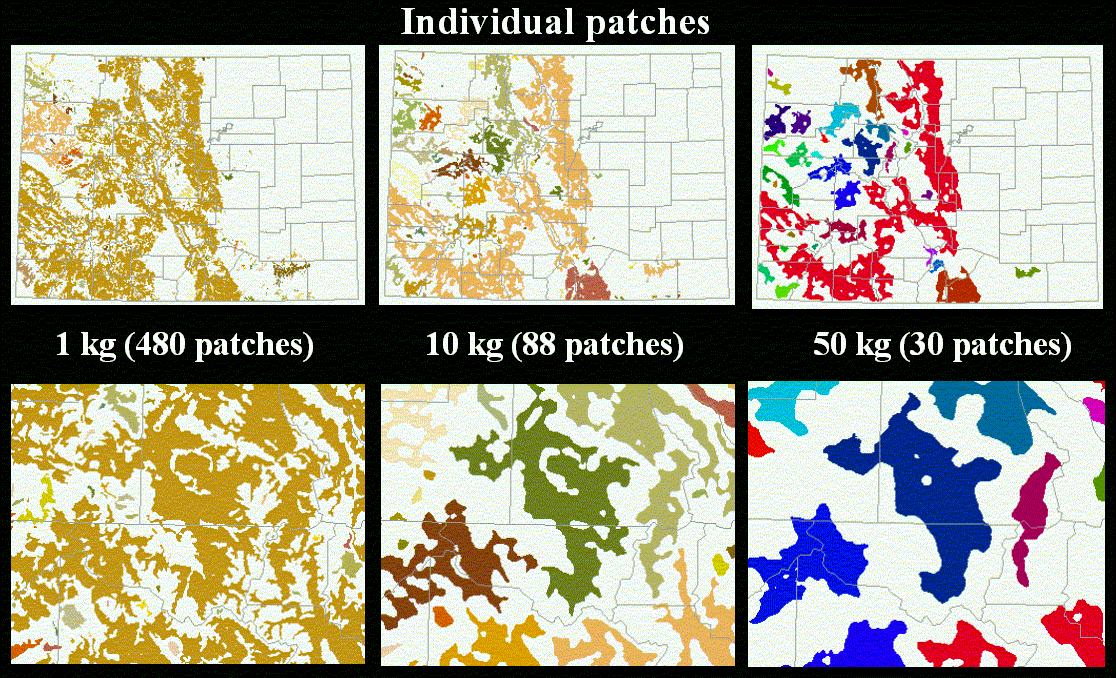
where M is body mass in kg and I is in km2. Next, we derive the radius required (RI) to fulfill the area requirement using Equation 3. For every cell in the habitat map we calculate the average habitat value within the radius RI using Equation 4. The map depicts the gradient of values that correspond to how likely a location is to contribute to an individual's habitat. To create the individual habitat patches, we threshold the gradient values, where the average quality PI in exceeds a user-specified efficiency threshold (t). Ultimately, the appropriate threshold value depends on the behavior of the species or process in consideration. We then aggregate adjacent (8 neighborhood) cells to form patches that contribute to individual's habitat (Figure 3).
RI = (I / 3.1415)1/2 (3)
PI = focalmean(Q, circle, RI ) > t (4)
Population
Population patches are identified in a similar fashion to the individual level, but we identify the area required to support minimum viable population patches based on allometric scaling from minimal mammal densities (Silva and Downing 1994) (Equations 5 and 6):
D = e(-0.68*ln(M)+2.1414) (5)
P = A / D (6)
where D is the minimum viable density in animals per km2 and P is the area required to support A animals.
Meta-population
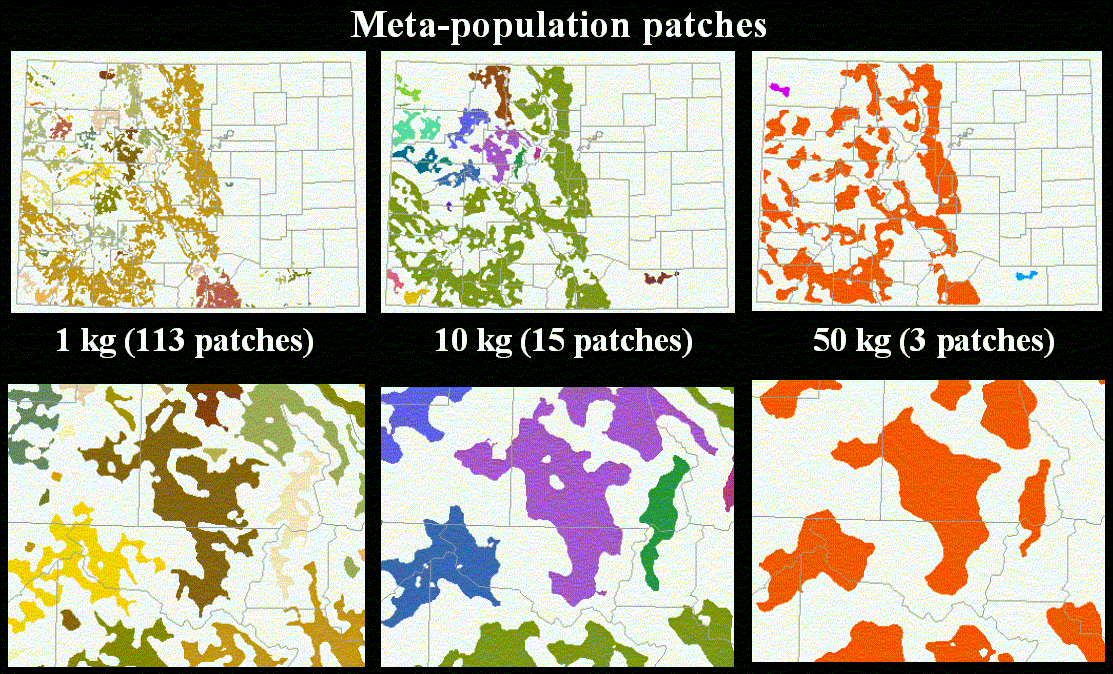
At a coarse-scale, movement is characterized by dispersal, and here the goal is to identify clusters of population patches that are within the dispersal distance of one another. Conversely, population patches are considered isolated when they are beyond the dispersal distance. As discussed above, the typical approach is to consider the dispersal the Euclidean distance from one patch to another (e.g., Keitt et al. 1997). But dispersal is affected not only by the location of habitat patches, but also the characteristics of the intervening patches that make up the matrix (Figure 4). The relative ease or difficulty a species has in moving through the matrix is largely influenced by the vegetation type at any given location. We conceptualize the matrix in terms of patches with impedance to movement and represent it using a cost surface, which allows Euclidean distance to be modified by the relative ease or difficulty to travel from one location to another. Also, human land use and natural disturbance regimes in the matrix interact with vegetation and can further restrict large-scale movement.
Again, we use allometric scaling to parameterize dispersal ability based on body size:
β
= 0.001M-0.91 (7)L = ln Z / - β (8)
where β is the probability of successful dispersal (Malkinson and Hobbs, unpublished), and L is the dispersal distance (m) for a probability of Z. We then calculate the distance based on matrix quality using a cost-distance function, where the population patches are the "source" patches for the cost-distance function. Typically, we reclassify land use/cover maps to reflect how the cover types would impede (or not) the movement through that area. The impedance values could also reflect other data layers, such as road or housing density, and very high values could cause a "barrier" to movement. Population patches are then grouped if they are within the distance L.
Example landscape
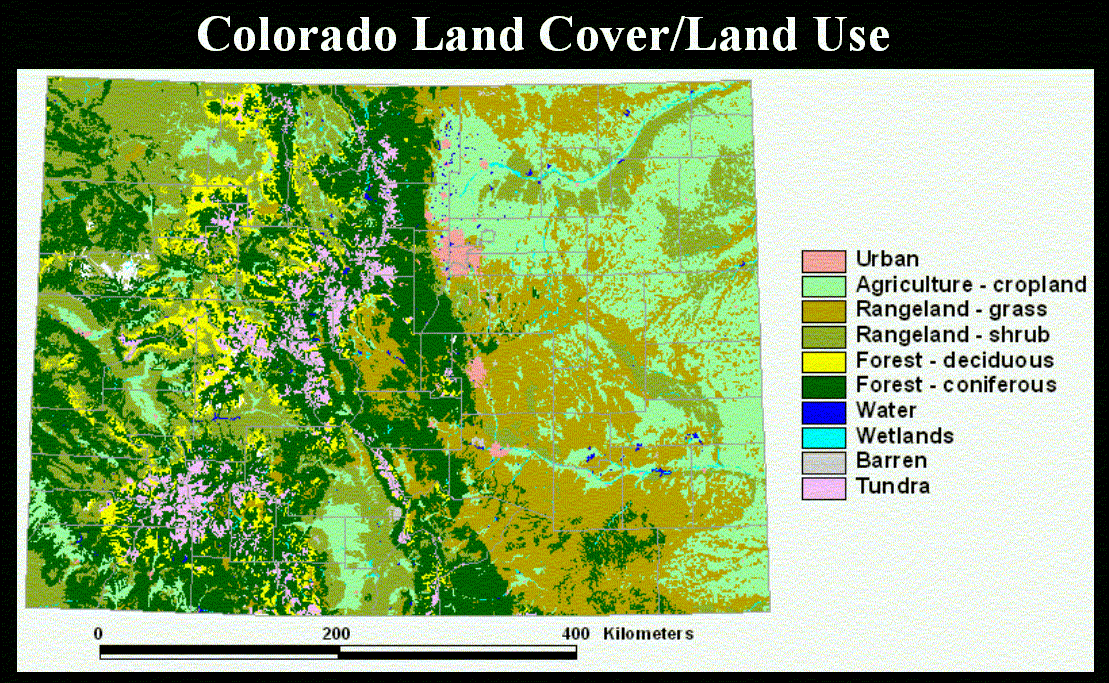
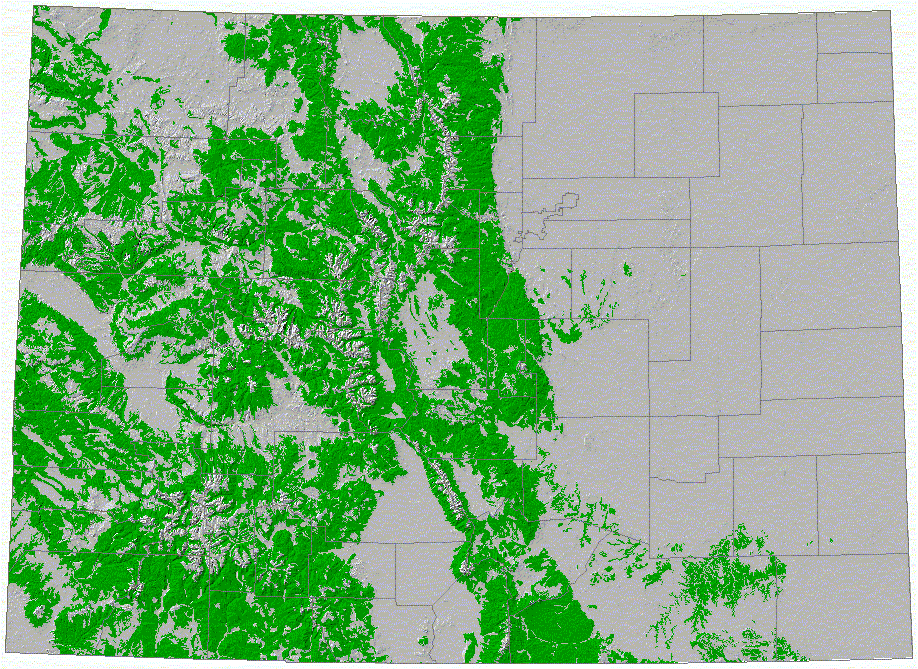
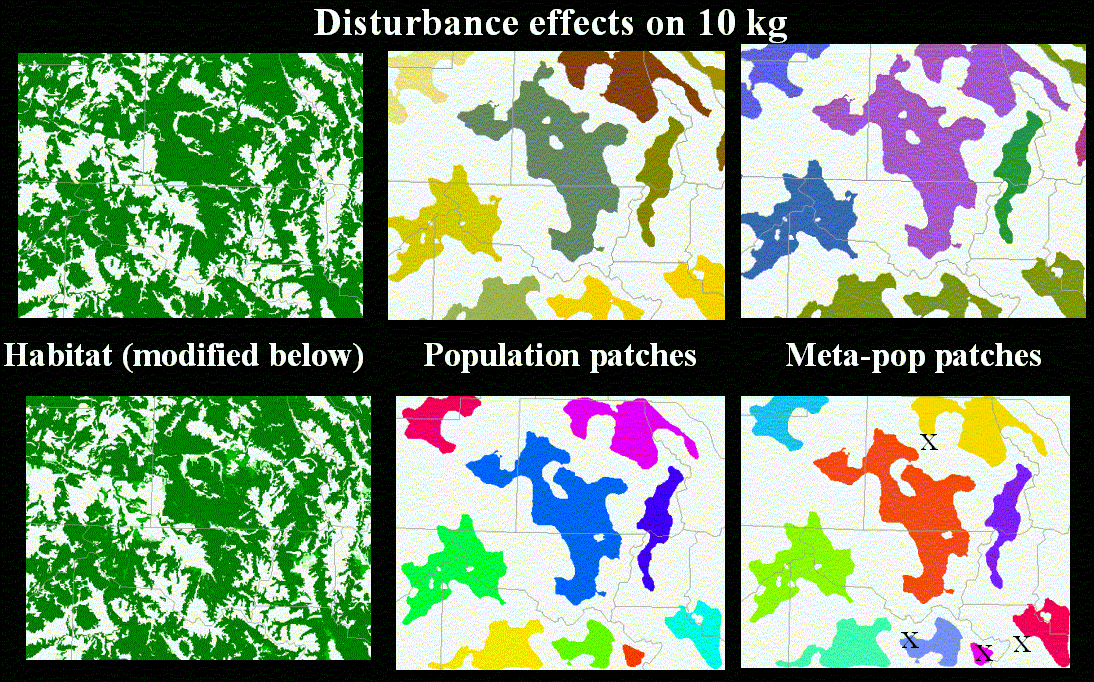
We illustrate our methodology by examining individual, population, and meta-population patches defined for a representative range of mammals (Table 1) in Colorado. The vegetation map we use from the Gap Analysis Project, which was produced by interpretation (using 100 ha minimum mapping unit) from Landsat TM image (30 m resolution) (Figure 5). We illustrate our approach by identifying forested-type habitats (including coniferous, deciduous, and mixed) (Figure 6). We then found the individual (Figure 7) and population (Figure 8) patches for mammals of 1, 10, and 50 kg in size and then reclassified the land use/cover map to reflect matrix quality and its influence on the inter-population-patch movement. We then calculated the meta-population patches (Figure 9), and Figure 10 shows meta-population patches after modifying the vegetation to simulate disturbance from housing development that reduces habitat quality.
Conclusions
Assessments that examine the consequences of development and land use change on habitat quality and fragmentation need to reflect the functional response of a species or process, rather than being simply an artifact of the data and interpretation scale. This requires biological parameters to be incorporated, and we find that parameterization of these models using allometric scaling of body size for three levels of organization is a useful way to incorporate biological realism into modeling habitat fragmentation. We plan to apply this model to inform reserve design by examining if there are situations when the stability of a reserve network is more sensitive to change in matrix quality than change in patch area.
References
Addicot, J.F., J.M. Aho, M.F. Antolin, D.K. Padilla, J.S. Richardson, and D.A. Soluk, 1987. Ecological neighborhoods: scaling environmental patterns. OIKOS 49: 340-346.
Baskent, E.Z., and G.A. Jordan, 1995. Characterizing spatial structure of forest landscapes. Canadian Journal of Forest Research 25:1830-1849.
Caicco, S.L., J.M. Scott, B. Butterfield, B. Csuti, 1995. A gap analysis of the management status of the vegetation of Idaho (U.S.A). Conservation Biology 9(3): 498-511.
Chen, J., J.F. Franklin, T.A. Spies, 1992. Vegetation responses to edge environments and old-growth Douglas-fir forests. Ecological Applications 2: 387-396.
Collinge, S.K. 1996. Ecological consequences of habitat fragmentation: implications for landscape architecture and planning. Landscape and Urban Planning 36: 59-77.
Congalton, R., 1991. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sensing of Environment 37: 35-46.
Dramstad, W.E., J.D. Olson, and R.T.T. Forman, 1996. Landscape ecology principles in Landscape Architecture and land-use planning. Island Press, Washington, D.C.
Edwards, T.C., Jr., E.T. Deshler, D. Foster, G.G. Moisen, 1996. Adequacy of wildlife habitat relation models for estimating spatial distributions of terrestrial vertebrates. Conservation Biology 10(1): 263-270.
Flather, C.H., K.R. Wilson, D.J. Dean, and W.C. McComb, 1997. Indentifying gaps in conservation networks: of indicators and uncertainty in geographic-based analyses. Ecological Applications 7(2): 531-542.
Gaston, K.J. and T.M. Blackburn, 1996. Conservation implications of geographic range size-body size relationships. Conservation Biology 10(2): 638-646.
Goodchild, M. F. and Gopal, S., (eds): 1989. The Accuracy of Spatial Databases. London, Taylor & Francis.
Gustafson, E.J. 1998. Quantifying landscape spatial pattern: what is the state of the art? Ecosystems 1: 143-156.
Haines-Young, R., D.R Green, and S.H. Cousins, 1993. Landscape ecology and spatial information systems, In: Landscape ecology and GIS, Haines-Young, R., D.R. Green, and S.H. Cousins, eds. Taylor & Francis, London.
Hobbs, N.T. 1999. Responses of large herbivores to spatial heterogeneity in ecosystems. In: Nutritional Ecology of Herbivores: Proceedings of the Vth International Symposium on the Nutrition of Herbivores, H. G. Jung and G. C. Fahey eds. American Society of Animal SciencePages 97-129. Savory IL.
Johnston, C.A. 1998. Geographic information systems in ecology. Blackwell Science, Oxford, U.K.
Keitt, T.H., D.L. Urban, B.T. Milne, 1997. Detecting critical scales in fragmented landscapes. Conservation Ecology 1(1):4.
MacArthur, R.H. and E.O. Wilson, 1967. The theory of island biogeography. Princeton University Press, Princeton, N.J.
McCullough, D.R. 1996. Metapopulations and wildlife conservation (ed.). Island Press, Washington, D.C.
O'Neill, R. V., Krummel, J. R., Gardner, R. H., Sugihara, G., Jackson, B., DeAngelis, D. L., Milne, B. T., Turner, M. G., Zygmunt, B., Christensen, S. W., Dale, V. H. and Graham, R. L. 1988. Indices of Landscape Pattern. Landscape Ecology 1(3): 153-162.
Peters, R.H. 1987. The ecological implications of body size. Cambridge University Press. pgs 329.
Reed, R.A., J.J. Johson-Barnard, and W.L. Baker 1996. Contribution of roads to forest fragmentation in the Rocky Mountains. Conservation Biology 10: 1098-1106.
Schumaker, N.H. 1996. Using landscape indices to predict habitat connectivity. Ecology 77(4): 1210-1225.
Shenk, T.M., Hobbs, N.T. and Theobald, D.M. 1997. Estimating the accuracy of Landsat TM vegetation classifications using fuzzy logic: A case study (poster). The Wildlife Society 1997 Annual Conference, Snowmass, Colorado.
Theobald, D.M. In review. Correcting linear and perimeter measurement errors in raster-based data. Cartography and Geographic Information Systems.
Silva, M. and J.A. Downing. 1994. Allometric scaling of minimal mammal densities. Conservation Biology 8(3): 732-743.
Theobald, D.M., Miller, J.M. and Hobbs, N.T. 1997. Estimating the cumulative effects of development on wildlife habitat. Landscape and Urban Planning 39(1): 25-36.
White, D., P.G. Minotti, M.J. Barczak, J.C. Sifneos, K.E. Freemark, M.V. Santelmann, C.F. Steinitz, A.R. Kiester, and E.M. Preston, 1997. Assessing risks to biodiversity from future landscape change. Conservation Biology 11(2):1-13.
Wiens, J.A. 1994. Habitat fragmentation: island v landscape perspectives on bird conservation. IBIS 137: S97-S104.
Wiens, J.A. 1996. Wildlife in patchy environments: Metapopulations, mosaics, and management. In: Metapopulations and wildlife conservation, D.R. McCullough, ed. Island Press, Washington D.C.
Wiens, J.A. In press. The science and practice of landscape ecology. In Landscape Ecological Analysis: Issues and Applications, Klopatek, J.M. and R.H. Gardner, eds. Springer-Verlag, New York.
Wiens, J.A., N.C. Stenseth, B. Van Horne, and R.A. Ims, 1993. Ecological mechanisms and landscape ecology. OIKOS 66:369-380.
Table 1. Parameters for a range of body sizes based on allometric relationships for mammals.
|
Body Size |
Individual Patch |
Population Patch |
Meta-population dispersal |
||
|
(kg) |
Area (km2) |
Radius (m) |
Area (km2) |
Radius (m) |
Distance (m) at p=0.5 |
|
1 |
1.17 |
609 |
11.75 |
1934 |
693 |
|
10 |
13.39 |
2064 |
56.23 |
4231 |
5634 |
|
50 |
73.72 |
4844 |
168.00 |
7313 |
24372 |
|
100 |
153.71 |
6995 |
269.15 |
9256 |
45796 |
|
200 |
320.47 |
10100 |
431.22 |
11716 |
86052 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}